But I'm still not in the habit of putting semicolons after everything. Despite studying Java and Scheme in high school, I never got very good with them or did anything practical. Right now, I'd say my native tongue is Python. Maybe if I get into automation, one of these days I'll
Monday, October 18, 2010
Chasing down particles
I'm up late writing MATLAB code to analyze a whole load of particle tracking data. It's not exactly a fun process, but I can feel myself getting better at MATLAB. It's like the coding equivalent of eating all my asparagus. There are some great tricks you can do to slice and dice arrays with fancy indexing. But, ugh, the syntax for optional arguments makes me wince (and avoid using optional arguments, unfortunately).
But I'm still not in the habit of putting semicolons after everything. Despite studying Java and Scheme in high school, I never got very good with them or did anything practical. Right now, I'd say my native tongue is Python. Maybe if I get into automation, one of these days I'llclamber up on the big-programmer potty learn C/C++.
But I'm still not in the habit of putting semicolons after everything. Despite studying Java and Scheme in high school, I never got very good with them or did anything practical. Right now, I'd say my native tongue is Python. Maybe if I get into automation, one of these days I'll
Monday, October 11, 2010
Storycest Is Best
For my folk song class, I'm writing an analysis of the old Scottish ballad Kemp Owyne (specifically the version called Kempion; here are some related ballads*). The most interesting part, I find, is looking at the relationships between the different versions and between Kemp Owyne and other stories.
Discovery number one: no version of Kemp Owyne is 'complete' in the sense of containing all the plot elements found in any version of Kemp Owyne. Furthermore, the different versions ('subspecies'?) seem to differ not randomly but systematically in the elements they omit. The Kempion version leaves out the vast majority of the beginning, so for example, you never find out why the evil stepmother curses the princess (in fact, you don't even find out it was the evil stepmother's fault until the very end), whereas other versions start out with a thread where the stepmother is angry about not being called "the fairest of them all". It's hard for me to even imagine what it's like to only know one version of the story, since I'm in the privileged position of being able to read them all and cross-reference.
Discovery number two: there's a lot more swapping of story elements than I thought -- a lot more random ligation of characters into new situations. I suppose most folktales are fanfiction of other folktales, for some sense of 'fanfiction'. For example, Kemp Owyne is supposed to be identified with Ywain/Yvain from the Arthurian legends, aka the historical Owain mab Urien (and "Kemp" means 'hero' or 'champion'). Why? No reason. He's just a convenient fictional knightly hero type. But interestingly enough, there's an invocation to St. Mungo at the end of Kempion... and yet, Yvain is also supposedly St. Mungo's father! How did that happen?? Either there is massive storycest going on, or someone chose to invoke a saint who wasn't even born yet. (You have to admit that time-traveling Scots heroes and saints would be pretty awesome.)
It really is like evolution, complete with horizontal gene transfer. But also with revival and retelling and all sorts of processes that don't have obvious biological analogues. Fascinating!
[*] Those texts have some typos in them, but I wasn't about to type all of Kemp Owyne again, and I'm really fond of that whole website because it makes up in quantity what little it lacks in quality. (The typos are few enough that I feel like only scholars really ought to care about them.)
Discovery number one: no version of Kemp Owyne is 'complete' in the sense of containing all the plot elements found in any version of Kemp Owyne. Furthermore, the different versions ('subspecies'?) seem to differ not randomly but systematically in the elements they omit. The Kempion version leaves out the vast majority of the beginning, so for example, you never find out why the evil stepmother curses the princess (in fact, you don't even find out it was the evil stepmother's fault until the very end), whereas other versions start out with a thread where the stepmother is angry about not being called "the fairest of them all". It's hard for me to even imagine what it's like to only know one version of the story, since I'm in the privileged position of being able to read them all and cross-reference.
Discovery number two: there's a lot more swapping of story elements than I thought -- a lot more random ligation of characters into new situations. I suppose most folktales are fanfiction of other folktales, for some sense of 'fanfiction'. For example, Kemp Owyne is supposed to be identified with Ywain/Yvain from the Arthurian legends, aka the historical Owain mab Urien (and "Kemp" means 'hero' or 'champion'). Why? No reason. He's just a convenient fictional knightly hero type. But interestingly enough, there's an invocation to St. Mungo at the end of Kempion... and yet, Yvain is also supposedly St. Mungo's father! How did that happen?? Either there is massive storycest going on, or someone chose to invoke a saint who wasn't even born yet. (You have to admit that time-traveling Scots heroes and saints would be pretty awesome.)
It really is like evolution, complete with horizontal gene transfer. But also with revival and retelling and all sorts of processes that don't have obvious biological analogues. Fascinating!
[*] Those texts have some typos in them, but I wasn't about to type all of Kemp Owyne again, and I'm really fond of that whole website because it makes up in quantity what little it lacks in quality. (The typos are few enough that I feel like only scholars really ought to care about them.)
Sunday, September 26, 2010
Science poetry: Heredity, Thomas Hardy
I am the family face;
Flesh perishes, I live on,
Projecting trait and trace
Through time to times anon,
And leaping from place to place
Over oblivion.
The years-heired feature that can
In curve and voice and eye
Despise the human span
Of durance -- that is I;
The eternal thing in man,
That heeds no call to die
--Thomas Hardy [source]
I like the alliterations and internal-half-rhymes in the first stanza. They remind me of what little I know about Anglo-Saxon verse. (I read Beowulf once...)
Monday, September 13, 2010

Sunday, September 5, 2010
$18 can buy a lot of fun
(My living group is doing Rush right now, which means my life is basically eaten for the next two weeks, such that I feel no shame about writing a punt-tastic post like this.)
I will just note that $18 can buy more dry ice than twenty MIT students can play around with for three hours. Try it out some time. Make sure to bring soapy water, interestingly-shaped glass vessels, small white and colored lights/LEDs, coins, spoons, and hot water.
Try putting a little water in a spoon and then resting the spoon on top of a chunk of dry ice. You can see the water freeze right before your eyes. It's pretty cool.
I will just note that $18 can buy more dry ice than twenty MIT students can play around with for three hours. Try it out some time. Make sure to bring soapy water, interestingly-shaped glass vessels, small white and colored lights/LEDs, coins, spoons, and hot water.
Try putting a little water in a spoon and then resting the spoon on top of a chunk of dry ice. You can see the water freeze right before your eyes. It's pretty cool.
Sunday, August 29, 2010
What's wrong with the cartoon eukaryotic cell? [Unusual Cells pt. 1]
I'm starting a series of posts based on a class I taught about "Unusual Cells" for Splash. Eventually, each post will include links to all the others.
The point of this series is to understand the wild and crazy unusual cells that populate the world (and that populate us!). But we should first understand the usual cell... or, perhaps, we will find that there is no such thing as a usual cell.
Consider the cartoon eukaryotic cell that we all know and, presumably, love:
 Figure 1: Typical eukaryotic cell. I'll refrain from listing the organelles, in order to prevent yawning; if you're curious, check Wikipedia, where this image is from.
Figure 1: Typical eukaryotic cell. I'll refrain from listing the organelles, in order to prevent yawning; if you're curious, check Wikipedia, where this image is from.
So... what's wrong with this cell? Here's a sampling of some of the answers my Splash students have given me:
The answer I'm really looking for is this:
(To be fair, Splash students pretty much always get this one too, and it's a general case of the answers I gave above except for #3.)
There's nothing in this cell! Yes, the major organelles are there, but where is the cytoskeleton? It's just that tiny little fiber (#7), which you can barely even see. Real cells are just packed with stuff. Structural proteins, membrane proteins, highways of vesicles wandering to and fro...
Now, of course, this is kind of a necessary evil. We can't include all that stuff in cartoons of the cell that are supposed to show the major organelles, because it would just be a distraction. Visual noise. For example, check out this slice of a more realistic watercolor:
 Figure 2: You can see a bit of the Golgi (yellow stacks) in this picture. The geodesic-dome-looking thing is a protein framework that's making a vesicle bud out from the Golgi. [Source]
Figure 2: You can see a bit of the Golgi (yellow stacks) in this picture. The geodesic-dome-looking thing is a protein framework that's making a vesicle bud out from the Golgi. [Source]
Seriously, check out the whole thing. It's beautiful. If I could get a quality print of this I would hang it above my bed. But even this is far from showing everything. The empty space between all those blobs is filled with crazy amounts of ions, small molecules, and of course water.
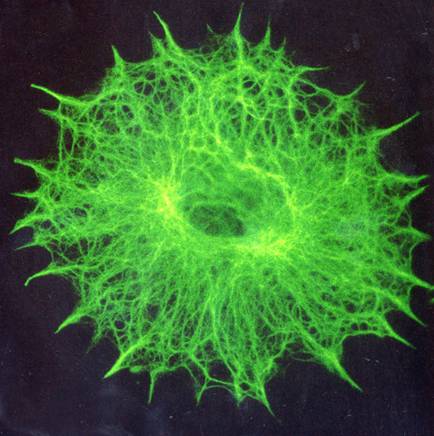
For a different perspective, check out these photos of cells in which the cytoskeleton has been labeled with green fluorescence. Yep, that's just the cytoskeleton... it reaches everywhere, helping the cell maintain its shape and move around (just like the human skeleton), and giving direction to packages of important chemicals as they motor their way hither and thither (something like the human circulatory system).

To do: find out what percentage of the membrane surface area is proteins. I know this is in one of my textbooks somewhere.
The point of this series is to understand the wild and crazy unusual cells that populate the world (and that populate us!). But we should first understand the usual cell... or, perhaps, we will find that there is no such thing as a usual cell.
Consider the cartoon eukaryotic cell that we all know and, presumably, love:
Figure 1: Typical eukaryotic cell. I'll refrain from listing the organelles, in order to prevent yawning; if you're curious, check Wikipedia, where this image is from.So... what's wrong with this cell? Here's a sampling of some of the answers my Splash students have given me:
- You can't see the DNA
- You can't see the proteins
- It's cut in half (yes, some of them are smartasses)
- There are no membrane proteins
The answer I'm really looking for is this:
It's empty!
(To be fair, Splash students pretty much always get this one too, and it's a general case of the answers I gave above except for #3.)
There's nothing in this cell! Yes, the major organelles are there, but where is the cytoskeleton? It's just that tiny little fiber (#7), which you can barely even see. Real cells are just packed with stuff. Structural proteins, membrane proteins, highways of vesicles wandering to and fro...
Now, of course, this is kind of a necessary evil. We can't include all that stuff in cartoons of the cell that are supposed to show the major organelles, because it would just be a distraction. Visual noise. For example, check out this slice of a more realistic watercolor:
Figure 2: You can see a bit of the Golgi (yellow stacks) in this picture. The geodesic-dome-looking thing is a protein framework that's making a vesicle bud out from the Golgi. [Source]Seriously, check out the whole thing. It's beautiful. If I could get a quality print of this I would hang it above my bed. But even this is far from showing everything. The empty space between all those blobs is filled with crazy amounts of ions, small molecules, and of course water.
For a different perspective, check out these photos of cells in which the cytoskeleton has been labeled with green fluorescence. Yep, that's just the cytoskeleton... it reaches everywhere, helping the cell maintain its shape and move around (just like the human skeleton), and giving direction to packages of important chemicals as they motor their way hither and thither (something like the human circulatory system).
To do: find out what percentage of the membrane surface area is proteins. I know this is in one of my textbooks somewhere.
Monday, August 23, 2010
Perhaps I should flip a coin?
I'm working out my class schedule for the upcoming semester, and I've run into a bit of a dilemma. Two classes are at the exact same time. Both clamor for my attention. Both are only offered in the fall, and this is my last year. They are as mutually exclusive as it is possible for two classes to be.
THE CANDIDATES
THE ARGUMENT
I want to study systems biology in grad school. Therefore, I should get started. Taking this class may help me with my continuing work in the Silver lab, and might even help me get into a good grad program.
THE COUNTERARGUMENT
I will have plenty of time to study systems biology in grad school. (And if I really get an itch, I can always pick up Uri Alon's book.) I should take this chance to explore a humanities topic that I'm really interested in, while I'm still an undergrad, because time is short.
------------------------------------------
So, what do I do? Both of these arguments are fairly convincing to me. Which one wins? Or, are there other arguments I've missed?
THE CANDIDATES
- 7.32 Systems Biology: This field is a sister to synthetic biology. I'm interested in to the point of wanting to pursue it in grad school. The Silver lab, where I'm working, is a systems biology lab in more than just name (although it's certainly not typical, being focused on engineering). Networks! Switches! Stochastic behavior! Dynamics! Oscillators! Pattern formation!
- 21A.212 Myth, Religion, and Symbolism: This class looks like it's going to hit one of my biggest avocational buttons. Despite being atheist/agnostic/nonreligious (damn labels), I've always had a fascination with the power of ritual and storytelling -- the roles they play in our lives and how they adapt to non-religious contexts. How did I manage to not notice this class existed before?
THE ARGUMENT
I want to study systems biology in grad school. Therefore, I should get started. Taking this class may help me with my continuing work in the Silver lab, and might even help me get into a good grad program.
THE COUNTERARGUMENT
I will have plenty of time to study systems biology in grad school. (And if I really get an itch, I can always pick up Uri Alon's book.) I should take this chance to explore a humanities topic that I'm really interested in, while I'm still an undergrad, because time is short.
------------------------------------------
So, what do I do? Both of these arguments are fairly convincing to me. Which one wins? Or, are there other arguments I've missed?
Sunday, August 22, 2010
Open thread: ask me questions!
Dear all the CS people who I know glance at this blog: I would love to hear from you! (And, y'know, from anyone else who happens not to study CS.)
I've recently come up short on blog topics, but also had a hankering to explain basic biology items in a way that makes them exciting to people who "hate biology", or are at best indifferent to it.
What would you like to hear about? Is there some old question left over from your intro biology course, to which you've never heard a satisfactory answer? Want quick summaries of recent developments in synthetic biology, or the ways in which biology imitates EE/CS? How about Anthropomorphized Enzyme Comics? Or, perhaps, White Lies Your High School Bio Teacher Told You?
Reply now and you might even get the post before Monday morning! :D
I've recently come up short on blog topics, but also had a hankering to explain basic biology items in a way that makes them exciting to people who "hate biology", or are at best indifferent to it.
What would you like to hear about? Is there some old question left over from your intro biology course, to which you've never heard a satisfactory answer? Want quick summaries of recent developments in synthetic biology, or the ways in which biology imitates EE/CS? How about Anthropomorphized Enzyme Comics? Or, perhaps, White Lies Your High School Bio Teacher Told You?
Reply now and you might even get the post before Monday morning! :D
Sunday, August 15, 2010
Science Poetry: The Perfume, A. D. Hope
I ran across this gem while trolling randomly through the archives of The Wondering Minstrels. It's the newest incarnation of the archives of an old mailing list, long gone out of service, but at least all the poems and commentary are still there. I'm in the process of going through all the poems, starting from no. 1. It's a wonderful archive, and I highly recommend spending some time there.
 Figure 1: Bombykol, from Wikipedia. "Doesn't that structure make you simply wild with desire?"
Figure 1: Bombykol, from Wikipedia. "Doesn't that structure make you simply wild with desire?"
I remember hearing stories about this compound, or one very like it, in my organic chemistry class. Apparently, whenever someone wanted to deliver a vial of it across campus, they would be pursued by a gradually accumulating swarm of moths. I like to play that scene in my head. "I'm a synthetic chemist -- I did not sign up for entomological fieldwork!! *panicked fleeing across campus*"
"... marked males of the silkworm moth have been known to fly upwind seven miles to a fragrant female of their kind ... the chemical compound with which a female silkworm moth attracts mates is highly specific; no other species seem aware of it. In 1959, the Nobel Laureate Adolph Butenandt of the Max Planck Institute for Biochemistry in Munich succeeded in analysing it. He found it to be an alcohol with sixteen carbon atoms per molecule...."
L. and M. Milne: The Senses of Animals and Men.
0 Chloë, have you heard it,
This news I sing to you?
It's true, my lovely bird, it
Is absolutely true!
A biochemist probing
Has caught without a doubt
The Queen of Love disrobing
And found her secret out.
What drives the Bombyx mori
To fly, intrepid male,
Lured by the old, old story
Six miles against the gale?
The formula, my Honey,
Is now in print to prove
What is, and no baloney,
The very stuff of love.
At Munich on the Isar
Those molecules were found
Which everyone agrees are
What makes the world go round;
What draws the male creation
To love, my darling doll,
Turns out, on trituration,
To be an alcohol.
A Nobel Laureatus
Called Adolph Butenandt
Contrived to isolate us
This strong intoxicant.
The boys are celebrating
And singing at the club:
Here's Bottoms up! to mating,
Since Venus keeps a pub!
My angel, 0, my angel,
What is it you suffuse,
What redolent evangel,
What nosegay of good news?
What draws me like a dragnet
And holds and keeps me tight?
What odds! my fragrant magnet,
I shall be drunk tonight!
-- A. D. Hope [source]
Figure 1: Bombykol, from Wikipedia. "Doesn't that structure make you simply wild with desire?"I remember hearing stories about this compound, or one very like it, in my organic chemistry class. Apparently, whenever someone wanted to deliver a vial of it across campus, they would be pursued by a gradually accumulating swarm of moths. I like to play that scene in my head. "I'm a synthetic chemist -- I did not sign up for entomological fieldwork!! *panicked fleeing across campus*"
Monday, August 9, 2010
Even supervillains have work-life balance problems
I just got back from seeing Despicable Me with my brother. Although this is not a Pixar movie, it follows Pixar's pattern by being about far, far more than the trailer lets on. (I remember being distinctly unimpressed by the WALL-E trailer, and then I cried my eyes out through the whole thing.) The plot is really fun, and the 3-D is unobtrusive enough to be mostly inconsequential.*
I thought it was pretty cool to see a supervillain (and an older, male supervillain at that) deal with the sort of struggles and discrimination normally associated with working mothers. I haven't experienced these struggles first-hand, of course, but a number of scenes reminded me very strongly of things I'd read -- especially Dr. Isis' blog posts and Allison Pearson's book I Don't Know How She Does It. Seeing parent-discrimination divorced from sexism was quite strange; but then again, I don't doubt that there are fathers in the world who have suffered career setbacks and discrimination because of family demands.
The end of the movie leaves open whether the main character continues the same level of career activity (in the same or a different field), or scales back in order to spend more time with the children. I would have liked to see something indicating that he achieved a productive balance; maybe a montage of newspaper headlines showing him up to something resembling his old tricks, perhaps with the children's collaboration.
Also, as a side note, whoever was writing Margo (the oldest girl) has done their homework on sibling-order effects on personality. I'm an oldest daughter, and though of course I'm not exactly like Margo I found myself identifying with her much more strongly than I was expecting to. (The girls in the movie are not biological siblings. Anyone know if sibling-order effects also happen in families brought together by adoption? It seems plausible -- these effects ought to be mediated in large part by environment... but I'm rambling now, because it's late.)
Go see Despicable Me if you're up for some funny, frivolous action, and so much cuteness that all your teeth will dissolve.
* I'm still cheesed at being made to pay an extra $4 for an effect that, IMO, adds very little to a movie animated in Pixar's style. But there's a cute bit in the credits where some minor characters play around with it, so stick around.
I thought it was pretty cool to see a supervillain (and an older, male supervillain at that) deal with the sort of struggles and discrimination normally associated with working mothers. I haven't experienced these struggles first-hand, of course, but a number of scenes reminded me very strongly of things I'd read -- especially Dr. Isis' blog posts and Allison Pearson's book I Don't Know How She Does It. Seeing parent-discrimination divorced from sexism was quite strange; but then again, I don't doubt that there are fathers in the world who have suffered career setbacks and discrimination because of family demands.
The end of the movie leaves open whether the main character continues the same level of career activity (in the same or a different field), or scales back in order to spend more time with the children. I would have liked to see something indicating that he achieved a productive balance; maybe a montage of newspaper headlines showing him up to something resembling his old tricks, perhaps with the children's collaboration.
Also, as a side note, whoever was writing Margo (the oldest girl) has done their homework on sibling-order effects on personality. I'm an oldest daughter, and though of course I'm not exactly like Margo I found myself identifying with her much more strongly than I was expecting to. (The girls in the movie are not biological siblings. Anyone know if sibling-order effects also happen in families brought together by adoption? It seems plausible -- these effects ought to be mediated in large part by environment... but I'm rambling now, because it's late.)
Go see Despicable Me if you're up for some funny, frivolous action, and so much cuteness that all your teeth will dissolve.
* I'm still cheesed at being made to pay an extra $4 for an effect that, IMO, adds very little to a movie animated in Pixar's style. But there's a cute bit in the credits where some minor characters play around with it, so stick around.
Wednesday, August 4, 2010
Gunnerkrigg Court takes on animal research ethics
I think I chose a particularly fortuitous time to highlight Gunnerkrigg Court, because it's just started taking on one of my favorite themes, and I think it's being handled very well so far.
First, read the last two pages of the comic: one, two. You don't need much context to see what I'm talking about.
 Figure 1: Rock on, Paz.
Figure 1: Rock on, Paz.
Now, if I remember correctly, Paz is a character we haven't seen much of yet (her first appearance is basically as an extra), and I'm looking forward to seeing her developed in more detail. I'm very glad to see she's taking (or at least professing) a sensitive, ethical attitude toward animal research. Realistic, too -- I'd swear that Tom Siddell has read the NIH Guidelines. I admire the fact that Paz aspires to reduce or even eliminate the use of animals in her research. Per fantasy conventions, all of these girls are stunningly mature and knowledgeable compared to the average high schooler, but I'm still very impressed by her attitudes and opinions.
(Plus, I'm pleased that she's apparently not white and not a native English speaker. I'm not terribly well informed about racial/identity politics, but nothing about the way she's portrayed jumps out at me as being problematic.)
I will be very interested to see how this plays out. In particular:
I have faith that all of these questions will be answered, if not in as much detail as I would like. Gunnerkrigg Court wouldn't just introduce a subplot like this without exploring it in quite a bit of detail.
First, read the last two pages of the comic: one, two. You don't need much context to see what I'm talking about.
 Figure 1: Rock on, Paz.
Figure 1: Rock on, Paz.Now, if I remember correctly, Paz is a character we haven't seen much of yet (her first appearance is basically as an extra), and I'm looking forward to seeing her developed in more detail. I'm very glad to see she's taking (or at least professing) a sensitive, ethical attitude toward animal research. Realistic, too -- I'd swear that Tom Siddell has read the NIH Guidelines. I admire the fact that Paz aspires to reduce or even eliminate the use of animals in her research. Per fantasy conventions, all of these girls are stunningly mature and knowledgeable compared to the average high schooler, but I'm still very impressed by her attitudes and opinions.
(Plus, I'm pleased that she's apparently not white and not a native English speaker. I'm not terribly well informed about racial/identity politics, but nothing about the way she's portrayed jumps out at me as being problematic.)
I will be very interested to see how this plays out. In particular:
- How will the teachers and other students at the Court react to the presence of animal research at their school? Will they even find out, or do they maybe know already? Will we see a range of attitudes, from "Animal welfare is not that important" to "All animal research is morally reprehensible"?
- How will the supernatural entities in the forest react? Will their reaction be shaped more by opposition to the Court in general, or by the fact that many of them are (at least in some sense) animals themselves?
- What is Paz doing, and who is she working with? How did she come by her research assistantship? Is she doing largely self-directed work or is she being used as a pawn by some unscrupulous adults? Is she aware of the broader implications of her work, whatever those turn out to be?
- Is this research actually justifiable/ethical or not? Right now all we have is Paz's word, and we have very little idea what they're actually studying.
I have faith that all of these questions will be answered, if not in as much detail as I would like. Gunnerkrigg Court wouldn't just introduce a subplot like this without exploring it in quite a bit of detail.
Friday, July 30, 2010
Watch out for the mad biologist!
One of my favorite webcomics is Gunnerkrigg Court. It's a mixed fantasy, SF, and school story with some characters who are pretty badass for their apparent age (middle/high school). Antimony and Kat, the two main characters, play a crucial, growing role in trying to preserve peace and increase cooperation between their school (the Court) and the adjacent forest -- but they're also just two girls capering in a world that's often a lot bigger than they realize. It's framed as technology versus magic/divinity, but I have a feeling it runs a lot deeper than that.
I highly recommend reading it, even though it starts out just a little slow -- the 'real' threads start soon enough.
But why am I highlighting it now? I'm super excited, because in among all the magicians and mad roboticists, I think we may have our first mad biologist character.
Should I say that again? Mad. Biologist. Character.
Squee!
Err, I would totally include a small click-to-embiggen preview image of the comic, but I can't figure out how to make Blogger do that. I have been thinking of migrating to Wordpress...
I highly recommend reading it, even though it starts out just a little slow -- the 'real' threads start soon enough.
But why am I highlighting it now? I'm super excited, because in among all the magicians and mad roboticists, I think we may have our first mad biologist character.
Should I say that again? Mad. Biologist. Character.
Squee!
Err, I would totally include a small click-to-embiggen preview image of the comic, but I can't figure out how to make Blogger do that. I have been thinking of migrating to Wordpress...
Monday, July 19, 2010
But why is the sky blue?
Here's a sampling of the random questions I've encountered in the past week that have piqued my interest. I feel like these should be answerable with a little effort (by someone other than me, since my brain is stuck in corners), but they're also fun to just speculate about.
It's a commonplace that Benadryl makes you drowsy. kcr noted that it only seems to make him drowsy when it isn't busy fighting off an actual allergic reaction. Are the antihistamine and sleep-inducing activities of Benadryl different? Does one compete with the other?
Why do martial arts seem to be optimized for fighting other practitioners of that same martial art? I don't know anything about how martial arts develop, but my intuition says something like this: it's dangerous and impractical to always practice by getting in real fights, and if you're going to make a new variant on an existing style, then practicing against others of that style is a decent proxy for being in real fights. Of course, this runs into the bootstrapping problem of where did the first formalized martial art come from... but humans have been punching each other for so many years that I feel like basic instinct can serve as a starting point. (How do you go about designing a martial art for "real combat"? I know these exist -- mumble military mumble something.)
What's the most cost-effective way to make ice cream using a dry ice and ethanol bath?
It's a commonplace that Benadryl makes you drowsy. kcr noted that it only seems to make him drowsy when it isn't busy fighting off an actual allergic reaction. Are the antihistamine and sleep-inducing activities of Benadryl different? Does one compete with the other?
Why do martial arts seem to be optimized for fighting other practitioners of that same martial art? I don't know anything about how martial arts develop, but my intuition says something like this: it's dangerous and impractical to always practice by getting in real fights, and if you're going to make a new variant on an existing style, then practicing against others of that style is a decent proxy for being in real fights. Of course, this runs into the bootstrapping problem of where did the first formalized martial art come from... but humans have been punching each other for so many years that I feel like basic instinct can serve as a starting point. (How do you go about designing a martial art for "real combat"? I know these exist -- mumble military mumble something.)
What's the most cost-effective way to make ice cream using a dry ice and ethanol bath?
Monday, July 12, 2010
A sure sign of a healthy lab
Whiteboard, day 1:
Whiteboard, day 2:
Paper attached to whiteboard, day 3:
Oh Danny boy, pipets, pipets are calling,
From bench to bench, and in the TC hood.
The gels are gone, and all the yields are falling;
'Tis you, 'tis you must make the data good.
So come ye back, where we cells are abiding,
Or when the lab is lonely as the grave.
'Tis here we'll be, in log-phase swift dividing...
...if you will please not put us in the autoclave!
THURS 3:30
GROUP MEETING
DANNY
GROUP MEETING
DANNY
Whiteboard, day 2:
THURS 3:30
GROUP MEETING
Oh DANNY boy
GROUP MEETING
Oh DANNY boy
Paper attached to whiteboard, day 3:
Oh Danny boy, pipets, pipets are calling,
From bench to bench, and in the TC hood.
The gels are gone, and all the yields are falling;
'Tis you, 'tis you must make the data good.
So come ye back, where we cells are abiding,
Or when the lab is lonely as the grave.
'Tis here we'll be, in log-phase swift dividing...
...if you will please not put us in the autoclave!
Friday, July 2, 2010
Bacteria break symmetry too
It's amazing what you can learn on the internet, especially when looking for something totally unrelated! :)
Caulobacter crescentus is a really cool little bacterial species with a funky two-phase lifestyle. The "stalked cells" attach themselves to rocks or whatever in the freshwater environments where these guys live. When a stalked cell divides, part of it remains a stalked cell and part splits off into a "swarmer cell". The swarmer cells swim around like more 'normal' bacteria.
Check out some images of these dudes. (Blogger's image uploader is misbehaving so you guys get a link to Google Images. Sigh.)
They're a fascinating organism to study because their cell division is asymmetrical. If you think back to the high-school-biology version of mitosis... well, it seems like a totally symmetrical process, right? There would seem to be no reason for a particular set of molecules to end up in one daughter cell and not the other, because everything's floating freely around in a droplet of water anyway. But in an asymmetric division like this, the two daughter cells have to develop in different ways. The stalked cell has to keep maintaining its stalk, but the swarmer cell has to grow a flagellum and start making the necessary sensory proteins to swim toward yummy-smelling food molecules. And not only that, but there's a correct orientation for this difference and an incorrect one. It would be kind of awkward if the new stalked cell started trying to swim away, and the swarmer cell floated around trying vainly to anchor to something.
So there must be some sophisticated mechanisms at play here. Notably, it's not that the two daughter cells end up with different genes -- after all, the swarmer cell will later settle and put down roots as a stalked cell. What matters is the presence (or absence) of proteins and other molecules that regulate those genes, so the stalked cell can keep making stalk proteins while the swimmer turns those genes off and turns on the ones for making a flagellum.
This asymmetric division isn't just some strange bacterial phenomenon. Every multicellular creature goes through this kind of process as it grows from a single cell (a fertilized egg) to whatever elaborate body it has as an adult. Figuring out the origins of symmetry-breaking in cell division is one of the major problems of developmental biology.
It also has to do with stem cells, by definition. The vast majority of cells in your body are "terminally differentiated" -- that is, they've gone from nondescript round blobs to fully elaborated cells with sophisticated morphology, heavily optimized for doing whatever job it is they need to do. But the 'terminally' part means they stop dividing once they reach maturity. So if you lose some mature cells, you need to get new ones from a renewable pool of immature cells. These are stem cells. The key defining feature of a stem cell is that it can divide asymmetrically. One of its progeny will be a precursor cell, traveling inexorably down the path to neuron-hood or white-blood-cell-hood or whatever. The other will be a new stem cell, all set to keep hanging in the lazy infinite loop of waiting until it's needed again.
Caulobacter crescentus is a really cool little bacterial species with a funky two-phase lifestyle. The "stalked cells" attach themselves to rocks or whatever in the freshwater environments where these guys live. When a stalked cell divides, part of it remains a stalked cell and part splits off into a "swarmer cell". The swarmer cells swim around like more 'normal' bacteria.
Check out some images of these dudes. (Blogger's image uploader is misbehaving so you guys get a link to Google Images. Sigh.)
They're a fascinating organism to study because their cell division is asymmetrical. If you think back to the high-school-biology version of mitosis... well, it seems like a totally symmetrical process, right? There would seem to be no reason for a particular set of molecules to end up in one daughter cell and not the other, because everything's floating freely around in a droplet of water anyway. But in an asymmetric division like this, the two daughter cells have to develop in different ways. The stalked cell has to keep maintaining its stalk, but the swarmer cell has to grow a flagellum and start making the necessary sensory proteins to swim toward yummy-smelling food molecules. And not only that, but there's a correct orientation for this difference and an incorrect one. It would be kind of awkward if the new stalked cell started trying to swim away, and the swarmer cell floated around trying vainly to anchor to something.
So there must be some sophisticated mechanisms at play here. Notably, it's not that the two daughter cells end up with different genes -- after all, the swarmer cell will later settle and put down roots as a stalked cell. What matters is the presence (or absence) of proteins and other molecules that regulate those genes, so the stalked cell can keep making stalk proteins while the swimmer turns those genes off and turns on the ones for making a flagellum.
This asymmetric division isn't just some strange bacterial phenomenon. Every multicellular creature goes through this kind of process as it grows from a single cell (a fertilized egg) to whatever elaborate body it has as an adult. Figuring out the origins of symmetry-breaking in cell division is one of the major problems of developmental biology.
It also has to do with stem cells, by definition. The vast majority of cells in your body are "terminally differentiated" -- that is, they've gone from nondescript round blobs to fully elaborated cells with sophisticated morphology, heavily optimized for doing whatever job it is they need to do. But the 'terminally' part means they stop dividing once they reach maturity. So if you lose some mature cells, you need to get new ones from a renewable pool of immature cells. These are stem cells. The key defining feature of a stem cell is that it can divide asymmetrically. One of its progeny will be a precursor cell, traveling inexorably down the path to neuron-hood or white-blood-cell-hood or whatever. The other will be a new stem cell, all set to keep hanging in the lazy infinite loop of waiting until it's needed again.
Thursday, June 24, 2010
Life lessons for synthetic biologists
1. A serious lesson
"Biology may or may not care about the physicist's insatiable desire for elegance." -- Jeff Hasty
 Figure 1: In other words, sometimes this happens. From Hasty et al, Physical Review Letters 2002 | doi:10.1103/PhysRevLett.88.148101
Figure 1: In other words, sometimes this happens. From Hasty et al, Physical Review Letters 2002 | doi:10.1103/PhysRevLett.88.148101
2. Another lesson that is just as serious
Humor in lab is essential, of course. However, if you just heard the great story about the giant biohazard bag full of innocuous things in your PI's car... finish laughing before you load your gel.
 The idea, of course, is to prevent this from happening. [Original photo source]
The idea, of course, is to prevent this from happening. [Original photo source]
"Biology may or may not care about the physicist's insatiable desire for elegance." -- Jeff Hasty
 Figure 1: In other words, sometimes this happens. From Hasty et al, Physical Review Letters 2002 | doi:10.1103/PhysRevLett.88.148101
Figure 1: In other words, sometimes this happens. From Hasty et al, Physical Review Letters 2002 | doi:10.1103/PhysRevLett.88.1481012. Another lesson that is just as serious
Humor in lab is essential, of course. However, if you just heard the great story about the giant biohazard bag full of innocuous things in your PI's car... finish laughing before you load your gel.
 The idea, of course, is to prevent this from happening. [Original photo source]
The idea, of course, is to prevent this from happening. [Original photo source]Sunday, June 13, 2010
MOSS!
I'm back from a 2.5-week family jaunt to Oregon and Washington. We bummed around a lot of national parks and did a wide variety of Wilderness Things. Here are some of the highlights:
Moss! We did go whale-watching, but for some reason I just don't get that excited about the large wildlife. I like to look at little things. And it turns out, the temperate rain forest is basically an ideal environment for dozens of moss species to thrive, reaching epic heights of lushness that I never dreamed of, growing up with miles of chaparral on all sides. There's lichens and liverworts too, but for some reason the mosses appeal to me the most. I discovered the macro setting on my camera (!) and took loads of pictures, trying to document as many different species as I could... but then I sat down too quickly on a rock and destroyed the screen. Argh! So I had to stop taking pictures. I don't even have the memory card with me, because my dad took it out when we got home and forgot to give it back to me before I flew back to Boston. So rest assured, I'll post my photos eventually. For now, here's a teaser photo taken by Derrick Ditchburn, who is a far better photographer than I. (More lovely moss photos at Dereila Images (do click on "More Moss" at the bottom).)
 Stair-step moss, the most elaborate moss I've ever seen. The main fronds get up to 3-4cm long, and grow in long dangling chains. Picture this carpeting an area of several meters square.
Stair-step moss, the most elaborate moss I've ever seen. The main fronds get up to 3-4cm long, and grow in long dangling chains. Picture this carpeting an area of several meters square.
Human-powered transport! I fell in love with biking and kayaking. I think the common thread is that they are both human-powered modes of transport that require a lot more thought than walking/running. I find them spectacularly engaging because I can pay attention to either the scenery or the vehicle, as I like. Plus, there's something satisfying about going twice or three times as fast as I could go unaided, but still without using a motor. I intend to continue both biking and kayaking in Boston -- I've borrowed a bike from a friend who's summering out of town, and apparently you can rent kayaks and go out on the Charles River.
Signing ghost! Yes, that said signing, not singing. We stopped in Ashland and caught a performance of Hamlet by the Oregon Shakespeare Festival, which was of course excellent. But one thing in particular struck me -- the ghost of Hamlet's father spoke in sign language! I looked through a couple of brochures and found out the ghost was played by Howie Seago, "the first deaf actor to play on OSF stages". I don't know any sign, so I couldn't tell whether he was using ASL or SEE or something else, but I thought it was a neat artistic choice to have him play the ghost, as opposed to a living character. (Hamlet spoke the ghost's lines, as if he only half-understood sign and was trying to keep up.) I bet translating Shakespeare into ASL is an interesting problem, too.
Moss! We did go whale-watching, but for some reason I just don't get that excited about the large wildlife. I like to look at little things. And it turns out, the temperate rain forest is basically an ideal environment for dozens of moss species to thrive, reaching epic heights of lushness that I never dreamed of, growing up with miles of chaparral on all sides. There's lichens and liverworts too, but for some reason the mosses appeal to me the most. I discovered the macro setting on my camera (!) and took loads of pictures, trying to document as many different species as I could... but then I sat down too quickly on a rock and destroyed the screen. Argh! So I had to stop taking pictures. I don't even have the memory card with me, because my dad took it out when we got home and forgot to give it back to me before I flew back to Boston. So rest assured, I'll post my photos eventually. For now, here's a teaser photo taken by Derrick Ditchburn, who is a far better photographer than I. (More lovely moss photos at Dereila Images (do click on "More Moss" at the bottom).)
Stair-step moss, the most elaborate moss I've ever seen. The main fronds get up to 3-4cm long, and grow in long dangling chains. Picture this carpeting an area of several meters square.Human-powered transport! I fell in love with biking and kayaking. I think the common thread is that they are both human-powered modes of transport that require a lot more thought than walking/running. I find them spectacularly engaging because I can pay attention to either the scenery or the vehicle, as I like. Plus, there's something satisfying about going twice or three times as fast as I could go unaided, but still without using a motor. I intend to continue both biking and kayaking in Boston -- I've borrowed a bike from a friend who's summering out of town, and apparently you can rent kayaks and go out on the Charles River.
Signing ghost! Yes, that said signing, not singing. We stopped in Ashland and caught a performance of Hamlet by the Oregon Shakespeare Festival, which was of course excellent. But one thing in particular struck me -- the ghost of Hamlet's father spoke in sign language! I looked through a couple of brochures and found out the ghost was played by Howie Seago, "the first deaf actor to play on OSF stages". I don't know any sign, so I couldn't tell whether he was using ASL or SEE or something else, but I thought it was a neat artistic choice to have him play the ghost, as opposed to a living character. (Hamlet spoke the ghost's lines, as if he only half-understood sign and was trying to keep up.) I bet translating Shakespeare into ASL is an interesting problem, too.
Monday, June 7, 2010
Wonder?
Christina Agapakis of Oscillator recently wrote a thought-provoking post about biosafety and synthetic biology. I was particularly struck by this passage:
I've often wondered myself why biology is so scary to so many people, and I think this is a particularly insightful way of putting it. If only more people would play around with sourdough starter or homemade yogurt, or manipulate soil pH to change the color of their hydrangeas, or hell just think harder about the fact that bacterial cells vastly outnumber human cells in a typical human body...
But of course, it's not that simple. Biology is hugely amazing or terrifying to a lot of people -- is there perhaps a good reason this is so? I feel like bio lab work has a really jading, mundanifying tendency: "I have the godlike power to manipulate the very genomes of bacteria!... and the result is that some of these spots are blue where none were blue before." The vast majority of the experiments I've personally done have ended either with a resounding "meh" or with a facepalm and a starting-over. Has this blinded me to the fact that, given sufficient equipment and time, I can engineer freaking life? How wondrous might genetic engineering seem to someone like Leeuwenhoek or Mendel?
What do y'all think? Especially you nonbiologists in the crowd? How weird does it seem to you that biologists collectively have these abilities? (And who's planning on seeing Splice?)
Why is biology scary to so many people? We've done a very good job of sterilizing our lives, separating ourselves from biology to the extant that when we think of the word bacteria we immediately think of infection that needs to be wiped out, not something that is part of our bodies, part of our everyday ecosystem that keeps us alive.
I've often wondered myself why biology is so scary to so many people, and I think this is a particularly insightful way of putting it. If only more people would play around with sourdough starter or homemade yogurt, or manipulate soil pH to change the color of their hydrangeas, or hell just think harder about the fact that bacterial cells vastly outnumber human cells in a typical human body...
But of course, it's not that simple. Biology is hugely amazing or terrifying to a lot of people -- is there perhaps a good reason this is so? I feel like bio lab work has a really jading, mundanifying tendency: "I have the godlike power to manipulate the very genomes of bacteria!... and the result is that some of these spots are blue where none were blue before." The vast majority of the experiments I've personally done have ended either with a resounding "meh" or with a facepalm and a starting-over. Has this blinded me to the fact that, given sufficient equipment and time, I can engineer freaking life? How wondrous might genetic engineering seem to someone like Leeuwenhoek or Mendel?
What do y'all think? Especially you nonbiologists in the crowd? How weird does it seem to you that biologists collectively have these abilities? (And who's planning on seeing Splice?)
Sunday, May 30, 2010
Reading the Histories of Middle-Earth
With characteristic good timing, I found myself suddenly interested in delving deeper into Tolkien's mythology... right around finals week. (Naturally, this led me to lie awake at night thinking about biological transport phenomena in the Two Trees of Valinor and so on.) But now I have a lot of free time and very little internet access, so I'm working my way through the first volume, The Book of Lost Tales I.
It takes a lot of getting used to. Everything's name is different, and the tales are interspersed with commentary from Christopher Tolkien, which is very insightful but detailed to the point of neuroticism. There are a number of subtle differences between this old material and the published Silmarillion, and very often I'm not sure which version I like better. (For example, in the Lost Tales, Ossë is a much more ambiguous character. He plays an important part in the story of the Lonely Isle, rather than just being "the Maia in charge of waves and storms".)
However, the big difference is that the whole history is framed as a series of tales told to a human mariner, Eriol, who comes to the Lonely Isle and hangs out with the Elves. And, well, it really doesn't work. Neither Eriol nor any of the tale-tellers is developed at all, although we are promised that actual plot events will happen later -- but they really need to happen closer to the beginning. The effect is basically "heroic adventurer arrives in distant mystical land and gets the Bible read to him for several weeks". It doesn't stand well on its own, but only in relation to LOTR, Hobbit, Silmarillion, etc. Let's just say I'm glad the stories were extensively revised before being published.
However, I'm only about halfway through the first history, and there are twelve, if I remember right. So I bet it gets interesting later. And after that, there's the Letters, where Tolkien gives a bunch more explicit commentary on the more philosophical ideas of his mythos -- mortality as Gift of Ilúvatar in particular. I'm looking forward to it!
It takes a lot of getting used to. Everything's name is different, and the tales are interspersed with commentary from Christopher Tolkien, which is very insightful but detailed to the point of neuroticism. There are a number of subtle differences between this old material and the published Silmarillion, and very often I'm not sure which version I like better. (For example, in the Lost Tales, Ossë is a much more ambiguous character. He plays an important part in the story of the Lonely Isle, rather than just being "the Maia in charge of waves and storms".)
However, the big difference is that the whole history is framed as a series of tales told to a human mariner, Eriol, who comes to the Lonely Isle and hangs out with the Elves. And, well, it really doesn't work. Neither Eriol nor any of the tale-tellers is developed at all, although we are promised that actual plot events will happen later -- but they really need to happen closer to the beginning. The effect is basically "heroic adventurer arrives in distant mystical land and gets the Bible read to him for several weeks". It doesn't stand well on its own, but only in relation to LOTR, Hobbit, Silmarillion, etc. Let's just say I'm glad the stories were extensively revised before being published.
However, I'm only about halfway through the first history, and there are twelve, if I remember right. So I bet it gets interesting later. And after that, there's the Letters, where Tolkien gives a bunch more explicit commentary on the more philosophical ideas of his mythos -- mortality as Gift of Ilúvatar in particular. I'm looking forward to it!
Thursday, May 20, 2010
Project idea: improving my attention span
By now it's a commonplace that the internet whittles your attention span down to the sort of tiny nub that only lets you focus on one thing for about thirty seconds before popping away to check email or RSS or Twitter or what have you. Word among my older MIT friends is that burnout and curricular exasperation can have similar effects. I'm inclined to believe them, having lost all patience with, say, biomechanics. *growl* *ahem* Where was I?
Oh, yes. I've been thinking that, among the other soul-restorative measures I plan to take this summer, I should work on reconstructing my attention span. According to my parents, when I was young I had a wonderful attention span, and could admire a pebble for ten minutes together. I'm not sure how much I believe them, but I do feel like my attention span has decreased dramatically over the past couple of years as I've started to read more and more things on the internet.
So I gave myself a test. I had to read something all the way through without looking at anything else on my computer. I picked the first interesting-looking article out of Give Me Something To Read, which turned out to be an NYTimes article about arranged remarriages in China following the 2008 Sichuan earthquake. (Incidentally, the article is very good -- thought-provoking and emotional without being overwrought.) Even though it was interesting, it was surprisingly difficult to get through. My mind kept wandering. Indeed, my mind is still wandering. I've pulled up my chat client once already while writing this blog post, and I keep having the urge to do it again. I'm like Randall Munroe at the typewriter here.
But with summer comes the opportunity to read books again! Do you have any idea how long it's been since I just sat down with a book and plowed more or less straight through the whole thing? I feel like I haven't done that since high school... So, as I pick up books again for the summer, I'll consciously practice keeping my attention on one thing for an extended period of time, and try to avoid get up every five minutes to get food or check my mail. We'll see how well I do, and whether this will translate to an improved studying efficiency in the fall.
Oh, yes. I've been thinking that, among the other soul-restorative measures I plan to take this summer, I should work on reconstructing my attention span. According to my parents, when I was young I had a wonderful attention span, and could admire a pebble for ten minutes together. I'm not sure how much I believe them, but I do feel like my attention span has decreased dramatically over the past couple of years as I've started to read more and more things on the internet.
So I gave myself a test. I had to read something all the way through without looking at anything else on my computer. I picked the first interesting-looking article out of Give Me Something To Read, which turned out to be an NYTimes article about arranged remarriages in China following the 2008 Sichuan earthquake. (Incidentally, the article is very good -- thought-provoking and emotional without being overwrought.) Even though it was interesting, it was surprisingly difficult to get through. My mind kept wandering. Indeed, my mind is still wandering. I've pulled up my chat client once already while writing this blog post, and I keep having the urge to do it again. I'm like Randall Munroe at the typewriter here.
But with summer comes the opportunity to read books again! Do you have any idea how long it's been since I just sat down with a book and plowed more or less straight through the whole thing? I feel like I haven't done that since high school... So, as I pick up books again for the summer, I'll consciously practice keeping my attention on one thing for an extended period of time, and try to avoid get up every five minutes to get food or check my mail. We'll see how well I do, and whether this will translate to an improved studying efficiency in the fall.
Monday, May 17, 2010
Silliness: Legos and BioBricks
It's finals week around here, which means you get a brief post while my brain tries to figure out electroosmosis and Van der Waals forces.
The similarities between Legos and BioBricks are legion -- in fact, if I'm explaining synthetic biology to someone and I've got more than ten minutes of their time, I'll break out my favorite Lego Analogies for the desirable properties that we want biological parts to have.
But they're similar in another way. Everyone refers to BioBricks as BioBricks, but you're supposed to call them BioBrick Standard Biological Parts.
 Trumpet fanfare optional. [Source]
Trumpet fanfare optional. [Source]
Likewise... hope I'm not ruining all your childhoods here... The LEGO(tm) corporation will be very, very sad if you refer to their product as "legos" instead of "LEGO bricks" or "LEGO toys". They might even cry.
 Apparently if you went to legos.com, you used to see this before being redirected, according to Mental Floss.
Apparently if you went to legos.com, you used to see this before being redirected, according to Mental Floss.
This sort of prescriptivism amuses me a little. I don't know enough about the relevant laws to comment on the legal necessity of being anal about plurals. But I was in a class co-taught by Drew Endy once, so I know he can say "BioBrick Standard Biological Parts" until he's blue in the face. Better him than me, I suppose.
The similarities between Legos and BioBricks are legion -- in fact, if I'm explaining synthetic biology to someone and I've got more than ten minutes of their time, I'll break out my favorite Lego Analogies for the desirable properties that we want biological parts to have.
But they're similar in another way. Everyone refers to BioBricks as BioBricks, but you're supposed to call them BioBrick Standard Biological Parts.
 Trumpet fanfare optional. [Source]
Trumpet fanfare optional. [Source]Likewise... hope I'm not ruining all your childhoods here... The LEGO(tm) corporation will be very, very sad if you refer to their product as "legos" instead of "LEGO bricks" or "LEGO toys". They might even cry.
 Apparently if you went to legos.com, you used to see this before being redirected, according to Mental Floss.
Apparently if you went to legos.com, you used to see this before being redirected, according to Mental Floss.This sort of prescriptivism amuses me a little. I don't know enough about the relevant laws to comment on the legal necessity of being anal about plurals. But I was in a class co-taught by Drew Endy once, so I know he can say "BioBrick Standard Biological Parts" until he's blue in the face. Better him than me, I suppose.
Sunday, May 9, 2010
Times when biology knowledge comes in useful
I love pineapple. Really, really love pineapple. Unfortunately, it irritates my mouth. I recently found out that this is not only due to the acidity -- pineapple contains a protease, bromelain. Bromelain will eat your face pretty effectively -- in fact, apparently there's a lot of interest in using it for wound debridement. [visceral shudder]
Unfortunately, the last time I ate a ton of pineapple all at once, I forgot about the protease until it was too late. But then I thought, "aha! I can saturate the protease with another type of protein and my mouth will remain unaffected!" Then I drank some milk.
Bio labs use milk as a generic solution of "loads and loads of proteins" in a lot of techniques. The one that springs to mind is Western blots. Basically, you run proteins through a gel that separates them by size and/or charge, to help identify what proteins you've got in the sample. Then you put your gel onto a nitrocellulose membrane that adsorbs proteins, so the spots from the gel transfer onto the membrane. Next, you want to probe the membrane with antibodies that should bind to your protein of interest, if it's on the membrane, and light up. But what's the problem? Antibodies are proteins, and the nitrocellulose membrane grabs onto all the proteins it touches, so unless you do something the antibody will just bind to the entire blot. What to do? Enter the milk! If you soak the membrane in milk before adding antibodies, then the milk proteins will bind all over the place and saturate the membrane, so then you can add antibodies without fear.
I told Zek about this and she mentioned another method for taking the bite out of pineapple: soak it in salt water. Apparently this is traditional in some places. We speculated that the high salt denatures the bromelain. I have yet to test whether this works or not, and whether it affects the taste.
Unfortunately, the last time I ate a ton of pineapple all at once, I forgot about the protease until it was too late. But then I thought, "aha! I can saturate the protease with another type of protein and my mouth will remain unaffected!" Then I drank some milk.
Bio labs use milk as a generic solution of "loads and loads of proteins" in a lot of techniques. The one that springs to mind is Western blots. Basically, you run proteins through a gel that separates them by size and/or charge, to help identify what proteins you've got in the sample. Then you put your gel onto a nitrocellulose membrane that adsorbs proteins, so the spots from the gel transfer onto the membrane. Next, you want to probe the membrane with antibodies that should bind to your protein of interest, if it's on the membrane, and light up. But what's the problem? Antibodies are proteins, and the nitrocellulose membrane grabs onto all the proteins it touches, so unless you do something the antibody will just bind to the entire blot. What to do? Enter the milk! If you soak the membrane in milk before adding antibodies, then the milk proteins will bind all over the place and saturate the membrane, so then you can add antibodies without fear.
I told Zek about this and she mentioned another method for taking the bite out of pineapple: soak it in salt water. Apparently this is traditional in some places. We speculated that the high salt denatures the bromelain. I have yet to test whether this works or not, and whether it affects the taste.
Monday, May 3, 2010
Meal planning: harder than it looks
Today was ET's Brunch in the Park, an event we throw every year for the current actives and alumni. A bunch of us made various breakfasty dishes. Watching the meal planning and preparation process was quite interesting. Most people went with something very simple, such as "pancakes" or "bacon", where the planning reduces to "Step 1: Buy a lot of whatever. Step 2: Cook all of it." I volunteered to make eggs, but I unwisely decided to get all fancy and make oven scrambled eggs with lots of mix-ins on the side.
My crucial mistake was not bothering to figure out the proper ratios -- I just went to the supermarket and bought "one" of everything: one head of broccoli, one 3-pack of bell peppers, etc. It turns out that one head of broccoli has significantly less mass than three bell peppers. It also turns out that it's hard to pan-fry things when all the stove space is continuously occupied by the pancake makers.
The worst part, though, was that the eggs were far too little and far too late. I cracked 25 eggs, but I should have done two or three times as many (or used an equivalent amount of that egg stuff that comes in cartons). We had around 30 people and only about 10 of them could have gotten an adequate amount of eggs. (As consolation, I can offer the fact that eggs + sauteed mushrooms + fresh basil = awesomeness!)
This episode gives me a whole new appreciation for the work our cook Karen does in planning meals and specifying what needs to be bought. She specifies the week's shopping list very neatly -- 4 pounds of broccoli, 3 cans of butter beans, foo units of bar, baz units of quux. Of course, she's had at least 20 years of practice, so I shouldn't be surprised that she's expert. After all, 2h/meal * 7 meals/wk * 20 full academic years works out to ~ 10,000 hours.
I just hope I can devote that kind of time and energy to some kind of lifetime pursuit or career...
My crucial mistake was not bothering to figure out the proper ratios -- I just went to the supermarket and bought "one" of everything: one head of broccoli, one 3-pack of bell peppers, etc. It turns out that one head of broccoli has significantly less mass than three bell peppers. It also turns out that it's hard to pan-fry things when all the stove space is continuously occupied by the pancake makers.
The worst part, though, was that the eggs were far too little and far too late. I cracked 25 eggs, but I should have done two or three times as many (or used an equivalent amount of that egg stuff that comes in cartons). We had around 30 people and only about 10 of them could have gotten an adequate amount of eggs. (As consolation, I can offer the fact that eggs + sauteed mushrooms + fresh basil = awesomeness!)
This episode gives me a whole new appreciation for the work our cook Karen does in planning meals and specifying what needs to be bought. She specifies the week's shopping list very neatly -- 4 pounds of broccoli, 3 cans of butter beans, foo units of bar, baz units of quux. Of course, she's had at least 20 years of practice, so I shouldn't be surprised that she's expert. After all, 2h/meal * 7 meals/wk * 20 full academic years works out to ~ 10,000 hours.
I just hope I can devote that kind of time and energy to some kind of lifetime pursuit or career...
Monday, April 26, 2010
Invasion of the possibly useful jargon
Every time I take a class, I semi-consciously pick up its jargon and use it for all kinds of unrelated things. I'm aware that this is very common among nerds/hackers -- after all, I hang out with lots of them. I, too, speak of "pinging" people in real life, and of the "failure modes" of couches and suchlike. But because I'm a biologist hanging out with mostly non-biologists, it stands out a lot more because everyone else isn't using jargon from their biochemistry classes. I don't know many other people who use words like "inhibit" and "saturation" and "depletion" and "steady state" and "modularity" on a daily-to-hourly basis. (OK, maybe the last two are more widespread than I think, and I just need to hang out with more MechE or EE people.)
The most recent one is "timescale", or "on the timescale of". I picked this one up from my biomechanics class, which analyzes bio-materials of all different sizes from single molecules to whole organs. In order to keep ourselves sane, we have to take into account the size of the object in question when choosing an analysis method. Should we speak of the stresses and strains in a rod made of continuous material, or of the entropy-driven behavior of a randomly meandering chain? Can we ignore thermal motion of molecules, or the transient behavior when you begin applying force? It all depends on the length scale.
I find the word "timescale" very useful in my daily life. It's much easier to say exactly what I mean if I say "on the timescale of weeks" rather than "in the medium-term". I would love to say that it helps other people understand, as well, but unfortunately no one else seems to have picked up on it yet, so I will have to wait and see.
Strangely enough, the jargon-adapting habit seems to be largely involuntary. However, the success or failure of a given word is definitely related to its usefulness, to the usefulness of the metaphor. There's not that much difference between a feedback system in a cafeteria and a piece of complex software, so it makes sense to speak of both of them as having "failure modes". I guess this is what the "seeing-as" theory of intelligence is all about. (Something I read in one of Hofstadter's books... I don't remember which it was, and I don't know what this is all about.)
Addendum: jargon-adapting is also not particularly widespread among people who aren't part of hacker or twinkie social circles. Or, at least, I don't encounter it very much, and I often get laughed at (in a kind way) when I'm hanging out with my friends from Bioengineering and I speak of rainwater "saturating" a drain, thus forming a puddle.
The most recent one is "timescale", or "on the timescale of". I picked this one up from my biomechanics class, which analyzes bio-materials of all different sizes from single molecules to whole organs. In order to keep ourselves sane, we have to take into account the size of the object in question when choosing an analysis method. Should we speak of the stresses and strains in a rod made of continuous material, or of the entropy-driven behavior of a randomly meandering chain? Can we ignore thermal motion of molecules, or the transient behavior when you begin applying force? It all depends on the length scale.
I find the word "timescale" very useful in my daily life. It's much easier to say exactly what I mean if I say "on the timescale of weeks" rather than "in the medium-term". I would love to say that it helps other people understand, as well, but unfortunately no one else seems to have picked up on it yet, so I will have to wait and see.
Strangely enough, the jargon-adapting habit seems to be largely involuntary. However, the success or failure of a given word is definitely related to its usefulness, to the usefulness of the metaphor. There's not that much difference between a feedback system in a cafeteria and a piece of complex software, so it makes sense to speak of both of them as having "failure modes". I guess this is what the "seeing-as" theory of intelligence is all about. (Something I read in one of Hofstadter's books... I don't remember which it was, and I don't know what this is all about.)
Addendum: jargon-adapting is also not particularly widespread among people who aren't part of hacker or twinkie social circles. Or, at least, I don't encounter it very much, and I often get laughed at (in a kind way) when I'm hanging out with my friends from Bioengineering and I speak of rainwater "saturating" a drain, thus forming a puddle.
Monday, April 19, 2010
Exhaustion
This semester, I've been mentoring a team of two freshmen working on designing a cooperative system of biofuel-producing algae and nutrient-recycling bacteria. It's a really neat project -- in fact all three of this year's 20.20 projects are really interesting. They have more of a focus on system dynamics / population engineering, where my year we focused on devices. I think this may be due to having Ron Weiss instead of Drew Endy. (Another indication of Weissitude is that we're all modeling our systems in MATLAB... sigh. I am not fond of MATLAB.)
I don't actually know much about biofuels, synthetic cooperation, or complex metabolism. I also don't know much about mentoring a team in the sort of hands-on, hands-off style that's appropriate when the point is for everyone to learn and stretch themselves. So it's a challenge, but an extremely satisfying one. I have watched my team drink the synthetic biology Kool-aid in the most remarkable way. There was a palpable transition from naivete to relative understanding;
Mentorship is important, but providing it at this level is also exhausting. Every day after class I feel physically tired (actually, that may have more to do with my sleep schedule than my mentoring schedule).
I don't actually know much about biofuels, synthetic cooperation, or complex metabolism. I also don't know much about mentoring a team in the sort of hands-on, hands-off style that's appropriate when the point is for everyone to learn and stretch themselves. So it's a challenge, but an extremely satisfying one. I have watched my team drink the synthetic biology Kool-aid in the most remarkable way. There was a palpable transition from naivete to relative understanding;
Mentorship is important, but providing it at this level is also exhausting. Every day after class I feel physically tired (actually, that may have more to do with my sleep schedule than my mentoring schedule).
Monday, April 5, 2010
Gaze Direction, or How Your RSI Prevention Software Can Distract Other People Too
The other day I noticed something funny happening. To set the scene, recall that humans are wired to detect other humans' gaze direction as a method of figuring out where the important stuff is happening at the moment. Robots are working on this too.
For the last several months I've been having problems with eye strain after using my computer for too long at a time, so I've been using RSI prevention software to remind me to look away from the screen every so often. (I use TimeOut (review here) because it doesn't reset its timer if you leave the mouse alone for 10 seconds while you're reading something.)
When anyone else is in the room and I look up from my laptop to stare fixedly at the opposite wall, about half the time they will look up too, wondering just what is so interesting about that light fixture or Periodic Table poster. It usually takes them several seconds to figure out that I'm not actually looking at anything in particular... we're just so used to thinking of gaze direction changes as highly meaningful.
Of course, this indicates to me that almost everyone I know needs to look away from their screens more often. Do it right now (seriously, I don't mind waiting) ...................................................................
...................................................................
...................................................................
...................................................................
...................................................................
...........................................................
you'll be surprised how your eyes feel when they refocus.
For the last several months I've been having problems with eye strain after using my computer for too long at a time, so I've been using RSI prevention software to remind me to look away from the screen every so often. (I use TimeOut (review here) because it doesn't reset its timer if you leave the mouse alone for 10 seconds while you're reading something.)
When anyone else is in the room and I look up from my laptop to stare fixedly at the opposite wall, about half the time they will look up too, wondering just what is so interesting about that light fixture or Periodic Table poster. It usually takes them several seconds to figure out that I'm not actually looking at anything in particular... we're just so used to thinking of gaze direction changes as highly meaningful.
Of course, this indicates to me that almost everyone I know needs to look away from their screens more often. Do it right now (seriously, I don't mind waiting) ...................................................................
...................................................................
...................................................................
...................................................................
...................................................................
...........................................................
you'll be surprised how your eyes feel when they refocus.
Monday, March 29, 2010
Overdue Ada Lovelace Day brief post
Typical. I don't get an idea for an Ada Lovelace Day post until well after the day itself has passed. "But I just don't know any women scientists other than the obvious ones!" Not an excuse. How many women scientists was I taught by in elementary and middle school, completely oblivious to their prior lives? Of course, I was taught by men scientists too, and they have equally interesting stories. I'm just going to focus on one particular woman for the moment.
Molly was the science specialist at my elementary school. She gave the impression of a powerful but friendly science warlock, introducing us grade-school apprentices to a world of wonderful things. Yes, it sounds cheesy, but there you are. I don't have any data, but I think I must credit Molly with giving me that initial (strong) inclination toward science that tipped and boiled over when I ran into a copy of The Cartoon Guide to Genetics (Gonick & Wheelis). It's been more than a decade since then, and I've never looked back.
I can only remember a few fragments of specific things that Molly taught us, but I distinctly remember some of the things that we did. My school had a lot of forested property around the buildings, some of which had been invaded by the vinca that someone decided to plant around the parking lots. We spent many an hour tramping around the woods, of course, and not only identifying trees and noticing erosion, but waging war on the vinca. We marked out test plots and poured mulch over some, staked black plastic over others, and wielded picks, shovels, and good ol' pulling on the rest. I lost track of the project once I was in middle school, so I don't know how successful the ongoing efforts were, but it was pretty awesome at the time.
We also -- this is kind of surprising, in retrospect -- had a strong focus on communication. At some level, every third grader is going to remember "communicating" (writing the occasional couple of paragraphs). But we wrote full-length letters to lawmakers about environmental issues (not skimping on either the science or the rhetoric), and drew a cartoon illustrating a forest fire. The final exhibition of everyone's cartoons was a sight to see.
The other day, I happened to idly wonder what Molly was up to now, and so I googled her. I found out little about what she's up to since her teaching career, but there was quite a bit about what she had been up to before taking up the overhead projector. She published several papers in the sixties about Blepharisma and the recovery of cells from X-ray irradiation. She was at Stanford! Doing real science! Back in the day, before sequencing and so on were invented! Why was I not informed?
She also had an active life outside school, running marathons and working with the San Francisco Sidewalk Astronomers to build DIY Dobsonian telescopes. Man, how cool is that?
Molly Lusignan, scientist, scholar, teacher, and all-around awesome woman, I salute you. The moral of the story for the rest of us is to find out what our teachers (and our children's teachers) do outside of the classroom, about their history, their hobbies, and their dreams. These days, it's of course made easy to stalk people via Google (and Google Scholar! <3), but there's a lot more to be said for connecting in person. I wish I could have found her current contact information...
Molly was the science specialist at my elementary school. She gave the impression of a powerful but friendly science warlock, introducing us grade-school apprentices to a world of wonderful things. Yes, it sounds cheesy, but there you are. I don't have any data, but I think I must credit Molly with giving me that initial (strong) inclination toward science that tipped and boiled over when I ran into a copy of The Cartoon Guide to Genetics (Gonick & Wheelis). It's been more than a decade since then, and I've never looked back.
I can only remember a few fragments of specific things that Molly taught us, but I distinctly remember some of the things that we did. My school had a lot of forested property around the buildings, some of which had been invaded by the vinca that someone decided to plant around the parking lots. We spent many an hour tramping around the woods, of course, and not only identifying trees and noticing erosion, but waging war on the vinca. We marked out test plots and poured mulch over some, staked black plastic over others, and wielded picks, shovels, and good ol' pulling on the rest. I lost track of the project once I was in middle school, so I don't know how successful the ongoing efforts were, but it was pretty awesome at the time.
We also -- this is kind of surprising, in retrospect -- had a strong focus on communication. At some level, every third grader is going to remember "communicating" (writing the occasional couple of paragraphs). But we wrote full-length letters to lawmakers about environmental issues (not skimping on either the science or the rhetoric), and drew a cartoon illustrating a forest fire. The final exhibition of everyone's cartoons was a sight to see.
The other day, I happened to idly wonder what Molly was up to now, and so I googled her. I found out little about what she's up to since her teaching career, but there was quite a bit about what she had been up to before taking up the overhead projector. She published several papers in the sixties about Blepharisma and the recovery of cells from X-ray irradiation. She was at Stanford! Doing real science! Back in the day, before sequencing and so on were invented! Why was I not informed?
She also had an active life outside school, running marathons and working with the San Francisco Sidewalk Astronomers to build DIY Dobsonian telescopes. Man, how cool is that?
Molly Lusignan, scientist, scholar, teacher, and all-around awesome woman, I salute you. The moral of the story for the rest of us is to find out what our teachers (and our children's teachers) do outside of the classroom, about their history, their hobbies, and their dreams. These days, it's of course made easy to stalk people via Google (and Google Scholar! <3), but there's a lot more to be said for connecting in person. I wish I could have found her current contact information...
Sunday, March 21, 2010
Journal Club Followup: The amber-suppressing AND gate
We went over the AND gate paper in class, and generated a lot of constructive criticism that I hadn't thought of by just reading it on my own. I guess this is what class is for, huh.
In particular, we addressed the question of whether the AND gate is truly modular. As I discussed in the previous post, in principle this AND gate is modular in the sense that it can be plugged into different inputs and outputs. However, this isn't the whole story. "Plugging it in" to a different input is not so simple as just putting a new promoter in front of Input Gene 1. The whole promoter/RBS/coding-region assembly has to be tuned to have an appropriate strength. If you swap in a stronger promoter, you might have to weaken the RBS (ribosome binding site) in order to get just the right amount of protein expression to feed in to the rest of the AND gate.
In fact, that's exactly what the authors had to do when they first put their AND gate together, which I kind of glossed over. Recall that the system works by expressing an mRNA with amber stop codons in, and the amber-suppressor tRNA that can read those stop codons; the mRNA codes for a protein that transcribes the output promoter. Now, in principle this should Just Work. But in practice, if there's enough mRNA around, you can get spontaneous read-through even if the amber-suppressor tRNA is supposedly turned "off", for two reasons. One, even if the tRNA is "off", it might still be produced at a low basal level. Two, the amber stop codon is relatively "weak", and sometimes just gets read through anyway. (That is, it's not very good at recruiting the translation-stopping machinery, which is supposed to disassemble the ribosome and cut loose the newly translated protein.) So the authors had to adjust the RBS that governed translation of the mRNA, so there wouldn't be too much of it floating around and you wouldn't get this spurious effect.
Apparently this sort of adjustment is called "impedance matching", for those of you who are more familiar with electronics than cells. I don't know enough about electronics to explain exactly what impedance matching is or why it's a suitable analogy here, but it boils down to "make sure all the wires are carrying appropriate amounts of current, and if you connect something new to a wire you might have to add a resistor or something to fix the current back to how it used to be."
So, no, it's not plug-and-play quite yet. More like plug, mutagenize the RBS, and play... but maybe we'll get there eventually.
The other main criticism of this AND gate, from a modularity point of view, is that you can't have two copies of the gate in the same cell and expect them to operate independently. mRNAs and tRNAs float around, and if one AND gate is expressing mRNA and the other is expressing tRNA, then both of them will output ON, even though both of them ought to be OFF. This is a more serious problem, because you can't just tune an RBS and expect it to go away -- this design for an AND gate is in principle not modular with respect to other AND gates placed in the same cell. (Possible workarounds include hiding different AND gates in different cells and mixing several populations together, but then you have to work with cell-to-cell cooperation, which is a whole different ballgame.)
In particular, we addressed the question of whether the AND gate is truly modular. As I discussed in the previous post, in principle this AND gate is modular in the sense that it can be plugged into different inputs and outputs. However, this isn't the whole story. "Plugging it in" to a different input is not so simple as just putting a new promoter in front of Input Gene 1. The whole promoter/RBS/coding-region assembly has to be tuned to have an appropriate strength. If you swap in a stronger promoter, you might have to weaken the RBS (ribosome binding site) in order to get just the right amount of protein expression to feed in to the rest of the AND gate.
In fact, that's exactly what the authors had to do when they first put their AND gate together, which I kind of glossed over. Recall that the system works by expressing an mRNA with amber stop codons in, and the amber-suppressor tRNA that can read those stop codons; the mRNA codes for a protein that transcribes the output promoter. Now, in principle this should Just Work. But in practice, if there's enough mRNA around, you can get spontaneous read-through even if the amber-suppressor tRNA is supposedly turned "off", for two reasons. One, even if the tRNA is "off", it might still be produced at a low basal level. Two, the amber stop codon is relatively "weak", and sometimes just gets read through anyway. (That is, it's not very good at recruiting the translation-stopping machinery, which is supposed to disassemble the ribosome and cut loose the newly translated protein.) So the authors had to adjust the RBS that governed translation of the mRNA, so there wouldn't be too much of it floating around and you wouldn't get this spurious effect.
Apparently this sort of adjustment is called "impedance matching", for those of you who are more familiar with electronics than cells. I don't know enough about electronics to explain exactly what impedance matching is or why it's a suitable analogy here, but it boils down to "make sure all the wires are carrying appropriate amounts of current, and if you connect something new to a wire you might have to add a resistor or something to fix the current back to how it used to be."
So, no, it's not plug-and-play quite yet. More like plug, mutagenize the RBS, and play... but maybe we'll get there eventually.
The other main criticism of this AND gate, from a modularity point of view, is that you can't have two copies of the gate in the same cell and expect them to operate independently. mRNAs and tRNAs float around, and if one AND gate is expressing mRNA and the other is expressing tRNA, then both of them will output ON, even though both of them ought to be OFF. This is a more serious problem, because you can't just tune an RBS and expect it to go away -- this design for an AND gate is in principle not modular with respect to other AND gates placed in the same cell. (Possible workarounds include hiding different AND gates in different cells and mixing several populations together, but then you have to work with cell-to-cell cooperation, which is a whole different ballgame.)
Tuesday, March 9, 2010
Where I'm going to be this summer
I hope you can forgive me for prematurely tooting my own horn, as there are many logistical details still to be worked out, but... I'm going to be working in Pam Silver's lab this summer! Squee!
 Figure 1: Yeast cell cycle counters! Hmm, those sound awfully familiar....
Figure 1: Yeast cell cycle counters! Hmm, those sound awfully familiar....
Figure 1: Yeast cell cycle counters! Hmm, those sound awfully familiar....Saturday, March 6, 2010
The answer to my question about scaffolds
I passed my scaffold question on to my 20.20 professors, and got a very thoughtful response. (To their credit, it was also a very timely response; I've just been slow in posting it.)
Recall my question: there seemed to be a conflict between the role of kinase cascades as signal amplifiers, and the role of scaffolds as giving specificity. Turns out, naturally, that there isn't really a conflict at all. Rather, I didn't fully understand what scaffolds do. The crux of what I wasn't understanding is that proteins go on and off their scaffold all the time -- they don't stay bound (semi)permanently.
One of the main benefits of scaffolds is to make signaling pathways more efficient: they help bring proteins close together so they can interact. If you have only a few molecules of Protein A and Protein B, then they may not encounter each other very often if they're just freely diffusing around the cell. A scaffold can grab one Protein A and one Protein B, hold them close together long enough to interact, and the release them and go look for two new proteins. Or if A is a kinase for B, then A can stay on the scaffold and multiple copies of B can take turns.
Scaffolds also act like little switchboards, dictating where signaling pathways are allowed to cross and merge or where they must stay separate. Suppose a scaffold has one binding site for the kinase A, and a second binding site that can fit either Protein B or C, but not D. In this case you get activation of both B and C, and the signal from A propagates in two different directions, provoking two separate responses (but not provoking a third response). This is really helpful because a lot of kinases are promiscuous -- they'll phosphorylate anything they can get their hands on -- so if they're confined to scaffolds, then they'll do whatever the scaffold says they can do, and no more.
Recall my question: there seemed to be a conflict between the role of kinase cascades as signal amplifiers, and the role of scaffolds as giving specificity. Turns out, naturally, that there isn't really a conflict at all. Rather, I didn't fully understand what scaffolds do. The crux of what I wasn't understanding is that proteins go on and off their scaffold all the time -- they don't stay bound (semi)permanently.
One of the main benefits of scaffolds is to make signaling pathways more efficient: they help bring proteins close together so they can interact. If you have only a few molecules of Protein A and Protein B, then they may not encounter each other very often if they're just freely diffusing around the cell. A scaffold can grab one Protein A and one Protein B, hold them close together long enough to interact, and the release them and go look for two new proteins. Or if A is a kinase for B, then A can stay on the scaffold and multiple copies of B can take turns.
Scaffolds also act like little switchboards, dictating where signaling pathways are allowed to cross and merge or where they must stay separate. Suppose a scaffold has one binding site for the kinase A, and a second binding site that can fit either Protein B or C, but not D. In this case you get activation of both B and C, and the signal from A propagates in two different directions, provoking two separate responses (but not provoking a third response). This is really helpful because a lot of kinases are promiscuous -- they'll phosphorylate anything they can get their hands on -- so if they're confined to scaffolds, then they'll do whatever the scaffold says they can do, and no more.
To see the beauty of a paper towel, and the world in an empty dewar
If you ever work in a bio lab, you'll find that often the closest piece of paper to hand is a paper towel, and the closest pen to hand is a sharpie or lab marker. Predictably, a lot of scientific note-taking happens on paper towels. I know I've done my share of calculations on paper towels, and I've heard of some students keeping their notebook as a stack of scribbled-on paper towels. (Shame!)
And then I walk down the infinite corridor and see something like this:
 Figure 1: Yes, Anonymous Lab Worker, someone appreciates your calligraphy. I love it when people do a little to make the world a weirder and more beautiful place. (Click to embiggen; the handwriting actually looks a lot better in high resolution.)
Figure 1: Yes, Anonymous Lab Worker, someone appreciates your calligraphy. I love it when people do a little to make the world a weirder and more beautiful place. (Click to embiggen; the handwriting actually looks a lot better in high resolution.)
And then I walk down the infinite corridor and see something like this:
Monday, March 1, 2010
Ideas dump
I'm looking over some old brainstorms, because of course I'm not busy at all, no sir, ahahaha... Anyway. I keep getting neat ideas, and I'm going to put some of them here so that (1) I don't forget them before I find time to investigate and (2) maybe someone will actually look at them and think about them. Also, I apologize for not explaining these ideas for the benefit of biology neophytes.
- Enzymes that bind DNA and perform actions including cutting/pasting DNA or recruiting other enzymes. Can you inhibit their action by adding short pieces of RNA (or similar) with the same sequence as their binding site?
- RNAi seems to exist in bacteria, kind of (not as much as in C.elegans). Is it useful for creating synthbiological devices? I don't recall seeing any; what's the roadblock?
- In particular, RNAi could be a neat way of getting around the crosstalk/specificity problem, since it's very sequence-specific and designing RNA sequences is easier than designing proteins.
- What would it take to drive localization to a synthetic organelle?
- Need to learn more about riboswitches. All the riboswitches I've seen so far are the kind that respond to a small molecule. Hmm: can you make a riboswitch that responds to a short ssRNA??
- What are the primitives of biological circuits? In a regime where it's easier to build monolithic black boxes than to reuse parts, how do things like two-component signaling evolve, that almost look intelligently designed with modularity etc?
- I think I'm on an RNA kick. Is this justified?
- Whoa, DNA scaffolds? Clever! read this at some point
Which tissues are anisotropic?
Today in my biomechanics class we moved out of molecular mechanics and into tissue mechanics. This will involve a lot of stereotypical mech-E stuff like stress and strain and elasticity that all us biologists have never heard of before -- deterministic, bulk properties of continuous solids (or gooey things), rather than stochastic models of single molecules or long chains of molecules being buffeted around by thermal motion.
In this field, as in every other, you have to make simplifying assumptions. Two of the key assumptions we often make about bulk materials are that they are homogeneous and isotropic. That is, they're uniform throughout, and they're the same in every direction. Tissue like muscle (with fibers) or substances like string cheese (also with fibers) are anisotropic (there's your five dollar word for the day). If you're stretching them and you want to find out how they deform, it matters which direction you're pulling. After we learned what all these fancy words meant, we categorized a few tissues:
Isotropic:
Anisotropic:
As I was writing these down, I noticed a pattern. All the isotropic tissues are what I'd call "biochemical tissues". Their main role is to store chemicals or make reactions happen. In contrast, all the isotropic tissues are "mechanical tissues", that make stuff move or stick together. Huh! Insight! But then it occurred to me that this makes total sense. After all, if you're doing a chemical reaction in a test tube, and you pour it into a bowl, the reaction will still happen. Whereas if you suddenly make all the fibers in a muscle run in a different direction, the muscle will do something completely different. If a tissue's job is to apply a force, it needs to apply that force in the right direction. Biochemical tissues are OK being isotropic, because their job is isotropic, and it seems like it ought to be harder for a developing organism to grow an anisotropic tissue, so why bother.
(Cartilage goes under "isotropic tissues", not because it does much in the way of chemical reactions, but because it's basically just a cushiony substance.)
Neat, huh?
Addendum: what about neurons and nerve tissue?
Well, we can throw out "neurons" straight out, because neurons are single cells, and tissues can only be treated as bulk materials if you've got a lot more than single cells. Trying to treat tissues on the tens-of-micrometers scale as bulk materials is like trying to calculate the viscosity of minestrone. It just doesn't apply. The viscosity of chicken broth is way different from the viscosity of beans. It only works if you zoom way, way out so that the effects of all the little bits and bobs become uniform over the entire blob of whatever you're looking at.
But we can totally look at nerve tissue this way. Your spinal cord, I would imagine, is a little bit like string cheese or rope. It's a whole bunch of long cellular fibers in parallel. If you pulled on it lengthwise, I would expect it to stretch; if you pulled on it widthwise, I would expect it to fray apart. It's anisotropic.
Gray matter in your cortex, on the other hand, I would call isotropic, or at least closer to isotropic (the cortex does have layers after all). Gray matter is mostly made up of cell bodies arranged more or less randomly, not a bunch of fibers all aligned with each other. Having dissected the odd brain or two, I think the best comparison for gray matter might be a firm jelly. (Exactly how firm it is depends on whether it's been preserved and how.)
So, does nerve tissue fit into the "biochemical" vs. "mechanical" tissue dichotomy? Not very cleanly. Then again, it's an atypical tissue. Its job is to send signals from point A to point B. For spinal cords and nerve bundles, point A and point B are far enough apart that it becomes important, on a macro scale, for the cells to go in the right direction. Gray matter contains mostly cell bodies rather than axons, so it's like a pile of Point As (or Points A, for the Captains Pedantic), whose job is to turn chemical signals into electrical activity.
In this field, as in every other, you have to make simplifying assumptions. Two of the key assumptions we often make about bulk materials are that they are homogeneous and isotropic. That is, they're uniform throughout, and they're the same in every direction. Tissue like muscle (with fibers) or substances like string cheese (also with fibers) are anisotropic (there's your five dollar word for the day). If you're stretching them and you want to find out how they deform, it matters which direction you're pulling. After we learned what all these fancy words meant, we categorized a few tissues:
Isotropic:
- Liver
- Fat
- Blood
- Cartilage
Anisotropic:
- Muscle
- Bone
- Skin
- Ligaments
As I was writing these down, I noticed a pattern. All the isotropic tissues are what I'd call "biochemical tissues". Their main role is to store chemicals or make reactions happen. In contrast, all the isotropic tissues are "mechanical tissues", that make stuff move or stick together. Huh! Insight! But then it occurred to me that this makes total sense. After all, if you're doing a chemical reaction in a test tube, and you pour it into a bowl, the reaction will still happen. Whereas if you suddenly make all the fibers in a muscle run in a different direction, the muscle will do something completely different. If a tissue's job is to apply a force, it needs to apply that force in the right direction. Biochemical tissues are OK being isotropic, because their job is isotropic, and it seems like it ought to be harder for a developing organism to grow an anisotropic tissue, so why bother.
(Cartilage goes under "isotropic tissues", not because it does much in the way of chemical reactions, but because it's basically just a cushiony substance.)
Neat, huh?
Addendum: what about neurons and nerve tissue?
Well, we can throw out "neurons" straight out, because neurons are single cells, and tissues can only be treated as bulk materials if you've got a lot more than single cells. Trying to treat tissues on the tens-of-micrometers scale as bulk materials is like trying to calculate the viscosity of minestrone. It just doesn't apply. The viscosity of chicken broth is way different from the viscosity of beans. It only works if you zoom way, way out so that the effects of all the little bits and bobs become uniform over the entire blob of whatever you're looking at.
But we can totally look at nerve tissue this way. Your spinal cord, I would imagine, is a little bit like string cheese or rope. It's a whole bunch of long cellular fibers in parallel. If you pulled on it lengthwise, I would expect it to stretch; if you pulled on it widthwise, I would expect it to fray apart. It's anisotropic.
Gray matter in your cortex, on the other hand, I would call isotropic, or at least closer to isotropic (the cortex does have layers after all). Gray matter is mostly made up of cell bodies arranged more or less randomly, not a bunch of fibers all aligned with each other. Having dissected the odd brain or two, I think the best comparison for gray matter might be a firm jelly. (Exactly how firm it is depends on whether it's been preserved and how.)
So, does nerve tissue fit into the "biochemical" vs. "mechanical" tissue dichotomy? Not very cleanly. Then again, it's an atypical tissue. Its job is to send signals from point A to point B. For spinal cords and nerve bundles, point A and point B are far enough apart that it becomes important, on a macro scale, for the cells to go in the right direction. Gray matter contains mostly cell bodies rather than axons, so it's like a pile of Point As (or Points A, for the Captains Pedantic), whose job is to turn chemical signals into electrical activity.
Thursday, February 25, 2010
Where do you learn?
Since gaining actual friends at college (I know, I know), I've been repeatedly surprised by how often people skip lecture. I must have noticed this earlier -- after all, I did observe that lecture halls tended to get less full as the semester went on -- but somehow I thought that all good students always went to lecture or some such. Apparently this is not the case.
Also, it seems like people are supposed to know by this time whether they're "visual learners" or "audio learners" or some such. I have never been able to figure this out. But apparently I learn pretty well by going to lectures (as opposed to doing the reading), although this is probably a motivation problem rather than anything else.
For 2.5 years this strategy served me well, but now that I run into courses with mediocre-to-poor lecturers who are always unclear and often omit large quantities of important material, it's becoming problematic.
Also, it seems like people are supposed to know by this time whether they're "visual learners" or "audio learners" or some such. I have never been able to figure this out. But apparently I learn pretty well by going to lectures (as opposed to doing the reading), although this is probably a motivation problem rather than anything else.
For 2.5 years this strategy served me well, but now that I run into courses with mediocre-to-poor lecturers who are always unclear and often omit large quantities of important material, it's becoming problematic.
Thursday, February 18, 2010
Learning to read all over again: Addendum
The project of learning to critically read the primary literature is going pretty well so far. I'd like to add another stage to the four stages I mentioned:
5. Selectivity. At this point you become able to read "the literature" as a whole, not just individual papers. You can make a good guess about what's important without having to read everything in excruciating detail.
I'll also note that the stages overlap quite a bit. I'm still mostly in Credulity, but there are some papers I can actually criticize, and some I still fail to understand.
5. Selectivity. At this point you become able to read "the literature" as a whole, not just individual papers. You can make a good guess about what's important without having to read everything in excruciating detail.
I'll also note that the stages overlap quite a bit. I'm still mostly in Credulity, but there are some papers I can actually criticize, and some I still fail to understand.
Wednesday, February 17, 2010
Impressions
The water dispenser outside my lab has a cold spigot and a hot spigot. The hot spigot dispenses water which is actually quite hot. However, I couldn't figure out how to use it at first... until I realized that you have to put the handle in a certain position, which is not its natural low-energy gravity-ordained position, and then press it. It's a safety interlock! Good thinking! *makes tea*
I drove in snow for the first time recently. Well, not snow, but there was some kind of sleet-like substance on the roads and falling (slowly) from the sky. It was also the first time I ever skidded (a small amount). I've been on a skid mat, but it was a while ago and I remember it very poorly. I think the problem with the skit mat was either that I was unable to actually skid, or that it made no sense out of the context of a street and the slight hazard of hitting parked cars. There's no spatial reference on an infinite frictionless plane, after all.
Plates of fermenting bacteria on a phenol-red containing medium have this wonderful tendency to grow into miniature sunrises (although they do develop a rather off-putting smell).
 Figure 1: Like viewing the sunrise from a mountaintop outhouse.
Figure 1: Like viewing the sunrise from a mountaintop outhouse.
I just reread Terry Pratchett's Going Postal, and I really liked it. He gives the sort of lovely far-off impressions of the world of the clacks towers that I want to go there but that might destroy the mystique -- you know, the same thing people always say about why the Silmarillion is written in such a dreamy mythic tone. And if Thief of Time is the Discworld's The Matrix, then Going Postal is its Jargon File.
I drove in snow for the first time recently. Well, not snow, but there was some kind of sleet-like substance on the roads and falling (slowly) from the sky. It was also the first time I ever skidded (a small amount). I've been on a skid mat, but it was a while ago and I remember it very poorly. I think the problem with the skit mat was either that I was unable to actually skid, or that it made no sense out of the context of a street and the slight hazard of hitting parked cars. There's no spatial reference on an infinite frictionless plane, after all.
Plates of fermenting bacteria on a phenol-red containing medium have this wonderful tendency to grow into miniature sunrises (although they do develop a rather off-putting smell).
 Figure 1: Like viewing the sunrise from a mountaintop outhouse.
Figure 1: Like viewing the sunrise from a mountaintop outhouse.I just reread Terry Pratchett's Going Postal, and I really liked it. He gives the sort of lovely far-off impressions of the world of the clacks towers that I want to go there but that might destroy the mystique -- you know, the same thing people always say about why the Silmarillion is written in such a dreamy mythic tone. And if Thief of Time is the Discworld's The Matrix, then Going Postal is its Jargon File.
Tuesday, February 16, 2010
Ruuuuuuuuuuuuuun!
I was reading my perfectly innocent-looking homework assignment, when all of a sudden I saw this:
 Figure 1: Augh!
Figure 1: Augh!
You know, I figured a biomechanics class (which is, after all, all about the realistic physicality of biology as opposed to magic cartoon enzymes that always work) would feature more realistic estimates of a bacterium's size.
 Figure 2: I also figured it would feature less fleeing and primal terror. [Source]
Figure 2: I also figured it would feature less fleeing and primal terror. [Source]
Of course, the answer is that E. coli are measured in μm (micrometers), not meters, and that the micro sign simply failed to render due to some strange failure of MS Word. The lesson is clear: Mistakes in character encodings will kill us all. Go forth boldly, my friends, and godspeed.
 Figure 1: Augh!
Figure 1: Augh!You know, I figured a biomechanics class (which is, after all, all about the realistic physicality of biology as opposed to magic cartoon enzymes that always work) would feature more realistic estimates of a bacterium's size.
 Figure 2: I also figured it would feature less fleeing and primal terror. [Source]
Figure 2: I also figured it would feature less fleeing and primal terror. [Source]Of course, the answer is that E. coli are measured in μm (micrometers), not meters, and that the micro sign simply failed to render due to some strange failure of MS Word. The lesson is clear: Mistakes in character encodings will kill us all. Go forth boldly, my friends, and godspeed.
Monday, February 15, 2010
Journal Club: The enzyme bucket brigade
JE Dueber, GC Wu, GR Malmirchegini, TS Moon, CJ Petzold, AV Ullal, KLJ Prather, & JD Keasling. Synthetic protein scaffolds provide modular control over metabolic flux. Nature Biotechnology 27, 753–759 (1 August 2009) | doi:10.1038/nbt.1557
I read and presented this paper in my lab class this past fall -- and I thought it was just one of the coolest papers I'd ever read. Cool concept, rational design, elegant solution to a Hard Problem, multiple benefits, modularity/composability, real-world results... this paper has it all.
You can look at cells as little factories, taking in raw materials and churning out interesting molecules. A cell's naturally occurring assembly lines are optimized by evolution in various ways, to increase efficiency and decrease interference ("cross-talk") with other processes in the cell. In particular, natural metabolic pathways are regulated so that they don't go wildly out of control and start overproducing whatever chemical, because that would be wasteful and expensive (not to mention potentially harmful).
But when you're putting an artificial assembly line into a cell, you have to undo some of these constraints and not others. You have to maximize efficiency, minimize cross-talk, and avoid making toxic products in the middle of the pathway as much as possible. These goals all line up with the goals of the cell. However, your main goal is different from the cell's goal of "produce just enough": you want to produce as much product as possible. More medicine. More biofuel. More super-protein-material-thing. More whatever. So this should be easy, right? The metabolic pathway is made up of enzymes that convert Chemical A to Chemical B to Chemical C, and you're inserting the genes for those enzymes into a bacterium. Why can't you just put very strong promoters in front of those genes, so you get massive quantities of each enzyme, and massive output?
This simple maxing-out approach causes several problems. First of all, it does nothing about the intermediate chemicals along the pathway -- they could still be toxic, or even just float away and go to waste. Second of all, this approach doesn't bother to optimize the ratio of the two enzymes. (If Enzyme 1 is half as efficient as Enzyme 2, then you ought to have twice as much of Enzyme 1.) Third of all, this doesn't do anything to stop the pathway cross-talking with other pathways. Fourth, and possibly most important, there's no guarantee that forcing each individual cell to make as much product as it possibly, possibly can is the most efficient way to convert cell food into useful chemicals. It's probably more efficient to let the cell divert plenty of energy into maintaining its own health and into spawning more cells, so you end up with more product overall.
To solve this dilemma, Dueber et al borrowed a trick that cells often use to regulate their own pathways. A scaffold is a structural protein that grabs on to all the enzymes in a given pathway, and holds them together into something like an assembly line or a bucket brigade.
I read and presented this paper in my lab class this past fall -- and I thought it was just one of the coolest papers I'd ever read. Cool concept, rational design, elegant solution to a Hard Problem, multiple benefits, modularity/composability, real-world results... this paper has it all.
You can look at cells as little factories, taking in raw materials and churning out interesting molecules. A cell's naturally occurring assembly lines are optimized by evolution in various ways, to increase efficiency and decrease interference ("cross-talk") with other processes in the cell. In particular, natural metabolic pathways are regulated so that they don't go wildly out of control and start overproducing whatever chemical, because that would be wasteful and expensive (not to mention potentially harmful).
But when you're putting an artificial assembly line into a cell, you have to undo some of these constraints and not others. You have to maximize efficiency, minimize cross-talk, and avoid making toxic products in the middle of the pathway as much as possible. These goals all line up with the goals of the cell. However, your main goal is different from the cell's goal of "produce just enough": you want to produce as much product as possible. More medicine. More biofuel. More super-protein-material-thing. More whatever. So this should be easy, right? The metabolic pathway is made up of enzymes that convert Chemical A to Chemical B to Chemical C, and you're inserting the genes for those enzymes into a bacterium. Why can't you just put very strong promoters in front of those genes, so you get massive quantities of each enzyme, and massive output?
This simple maxing-out approach causes several problems. First of all, it does nothing about the intermediate chemicals along the pathway -- they could still be toxic, or even just float away and go to waste. Second of all, this approach doesn't bother to optimize the ratio of the two enzymes. (If Enzyme 1 is half as efficient as Enzyme 2, then you ought to have twice as much of Enzyme 1.) Third of all, this doesn't do anything to stop the pathway cross-talking with other pathways. Fourth, and possibly most important, there's no guarantee that forcing each individual cell to make as much product as it possibly, possibly can is the most efficient way to convert cell food into useful chemicals. It's probably more efficient to let the cell divert plenty of energy into maintaining its own health and into spawning more cells, so you end up with more product overall.
To solve this dilemma, Dueber et al borrowed a trick that cells often use to regulate their own pathways. A scaffold is a structural protein that grabs on to all the enzymes in a given pathway, and holds them together into something like an assembly line or a bucket brigade.
A question about scaffolds
There are two bits of signal transduction dogma that have started to bother me. I don't know why I didn't spot this before.
Kinases are proteins that activate other proteins by attaching phosphate groups to them. Kinase cascades (several kinases in a row) are fairly common in all kinds of signaling pathways. Their main benefits are amplifying and diversifying the signal. Since a kinase is after all an enzyme, it can catalyze the same reaction over and over; it can activate many copies of the next kinase in the cascade, each of which can activate many copies of... and so on into exponential growth. That's how you get amplification, turning a tiny-but-important input into a massive cell-wide response. Diversification comes in when a kinase has more than one target. This comes in handy when the cell needs to respond to one signal by doing several different things all at once.
Scaffolds are large structural proteins that grab several other proteins from a signaling (or metabolic) pathway and hold them together. This helps them get their job done more efficiently. They help make pathways specific. If Enzyme 2 is stuck on a scaffold between Enzyme 1 and Enzyme 3, it can't very well run off to some other part of the cell and mess something up.
So, I perceive a slight conflict here. On one hand, it's helpful to diversify a signal; on the other hand, signals ought to be specific. On one hand, enzyme cascades amplify signals by working catalytically instead of stoichiometrically; on the other hand, when kinases are bound to a scaffold, their stoichiometric ratio is locked at one-to-one. What's going on here?
The obvious answer is that each type of signal processing is used where it's appropriate, and all types are appropriate in different contexts. If this is the explanation, then you would never expect to find a kinase cascade associated with a scaffold. But that's exactly what the MAP kinase cascade does! What gives? I thought the whole point of having a kinase cascade was to amplify and diversify the signal, which is exactly what the scaffold seems to be preventing.
 Figure 1: MAP kinase cascade shamelessly associating with scaffold protein. Have they no shame? What has the yeast
Figure 1: MAP kinase cascade shamelessly associating with scaffold protein. Have they no shame? What has the yeast mating pathway courting ritual come to these days? [Source]
Kinases are proteins that activate other proteins by attaching phosphate groups to them. Kinase cascades (several kinases in a row) are fairly common in all kinds of signaling pathways. Their main benefits are amplifying and diversifying the signal. Since a kinase is after all an enzyme, it can catalyze the same reaction over and over; it can activate many copies of the next kinase in the cascade, each of which can activate many copies of... and so on into exponential growth. That's how you get amplification, turning a tiny-but-important input into a massive cell-wide response. Diversification comes in when a kinase has more than one target. This comes in handy when the cell needs to respond to one signal by doing several different things all at once.
Scaffolds are large structural proteins that grab several other proteins from a signaling (or metabolic) pathway and hold them together. This helps them get their job done more efficiently. They help make pathways specific. If Enzyme 2 is stuck on a scaffold between Enzyme 1 and Enzyme 3, it can't very well run off to some other part of the cell and mess something up.
So, I perceive a slight conflict here. On one hand, it's helpful to diversify a signal; on the other hand, signals ought to be specific. On one hand, enzyme cascades amplify signals by working catalytically instead of stoichiometrically; on the other hand, when kinases are bound to a scaffold, their stoichiometric ratio is locked at one-to-one. What's going on here?
The obvious answer is that each type of signal processing is used where it's appropriate, and all types are appropriate in different contexts. If this is the explanation, then you would never expect to find a kinase cascade associated with a scaffold. But that's exactly what the MAP kinase cascade does! What gives? I thought the whole point of having a kinase cascade was to amplify and diversify the signal, which is exactly what the scaffold seems to be preventing.
 Figure 1: MAP kinase cascade shamelessly associating with scaffold protein. Have they no shame? What has the yeast
Figure 1: MAP kinase cascade shamelessly associating with scaffold protein. Have they no shame? What has the yeast Journal Club: The amber-suppressing AND gate
Anderson JC, Voigt CA, Arkin AP. Environmental signal integration by a modular AND gate. Molecular Systems Biology 3:133 (2007) | doi:10.1038/msb4100173
Logic gates (AND, OR, NOT, etc.) are the basis of electronic computation. If we'd like to implement biological computation, one of our first steps has to be implementing similar logic gates using proteins and DNA. That is, we need to make devices that accept a few inputs, perform a logical operation on them, and then spit out the result. In the case of an AND gate, we want the output to be ON whenever both of the inputs are ON, and OFF when either input is OFF. It seems easy, but of course, this turns out to be a lot harder in biology (hence, people writing papers about it).
 Figure 1: The result of this paper, if you abstract away all the interesting stuff. [Source]
Figure 1: The result of this paper, if you abstract away all the interesting stuff. [Source]
What makes it hard to make an AND gate out of biochemical parts?
Lack of standard connectors. In electronics, every signal is carried by a current, and every connector is a wire. That isn't the case in biology. Biological signals are typically carried by the presence or absence of some protein that carries out some particular chemical reaction that affects other proteins. This is wildly nonstandardized, and it means if Protein A interacts with Receptor A, you can't just plug in Receptor B and expect things to work.
Fuzzy, non-discrete behavior. It's nearly impossible for a biological system to have a perfect ON or OFF state. Even if a signal is mostly off, there'll be a few molecules of it floating around somewhere. And when you go to turn it on, it'll take time. Basically, biological things tend to vary continuously and not discretely (in large jumps).
Crosstalk. If a biological device relies on some particular molecule, then that molecule is going to be everywhere in the cell. So, you can't put two copies of the same device into a cell and expect them to operate independently. They'll interfere or "crosstalk" with each other in ways you don't expect. In contrast, you can throw down dozens of electronic circuit elements onto a breadboard and they won't interfere with each other because they're separated by physical space. In biology, everything's in the same soup.
Logic gates (AND, OR, NOT, etc.) are the basis of electronic computation. If we'd like to implement biological computation, one of our first steps has to be implementing similar logic gates using proteins and DNA. That is, we need to make devices that accept a few inputs, perform a logical operation on them, and then spit out the result. In the case of an AND gate, we want the output to be ON whenever both of the inputs are ON, and OFF when either input is OFF. It seems easy, but of course, this turns out to be a lot harder in biology (hence, people writing papers about it).
Figure 1: The result of this paper, if you abstract away all the interesting stuff. [Source]What makes it hard to make an AND gate out of biochemical parts?
Lack of standard connectors. In electronics, every signal is carried by a current, and every connector is a wire. That isn't the case in biology. Biological signals are typically carried by the presence or absence of some protein that carries out some particular chemical reaction that affects other proteins. This is wildly nonstandardized, and it means if Protein A interacts with Receptor A, you can't just plug in Receptor B and expect things to work.
Fuzzy, non-discrete behavior. It's nearly impossible for a biological system to have a perfect ON or OFF state. Even if a signal is mostly off, there'll be a few molecules of it floating around somewhere. And when you go to turn it on, it'll take time. Basically, biological things tend to vary continuously and not discretely (in large jumps).
Crosstalk. If a biological device relies on some particular molecule, then that molecule is going to be everywhere in the cell. So, you can't put two copies of the same device into a cell and expect them to operate independently. They'll interfere or "crosstalk" with each other in ways you don't expect. In contrast, you can throw down dozens of electronic circuit elements onto a breadboard and they won't interfere with each other because they're separated by physical space. In biology, everything's in the same soup.
Tuesday, February 9, 2010
Learning to read all over again
In my RNAi seminar, one of the instructors (they're both postdocs) noted that reading primary literature is a legitimately difficult thing. He expects us all to progress through several stages:
I think I'm more or less at the Credulity stage. I can figure out what a paper is about, though it takes me a while, and I get super excited about the key results. I've gotten to the point where I can read papers by myself, but I've never seen a paper picked apart, analyzed, and criticized in any detail where I could actually follow the conversation (this is hard in high school). So I think I'll be needing professional help, as it were, to move beyond Credulity.
(I might have gone a little bit overboard with the paper-reading classes for this semester, though. I added it up and I think I'm going to be reading 6-8 papers a week, or about one per day. Wow.)
- Incomprehension. At first, it's difficult to even figure out what the authors are talking about, or trace the flow of experimental logic. Sometimes it just doesn't flow quite right; other times you get distracted by all the technical details you don't understand at all or don't understand the need for.
- Credulity. Once you learn how a few procedures work and start to understand why people do experiments in the order they do, you can finally pull out a paper's main substantive point from all the noise. At this point you'll believe whatever the paper says, because the graphs look pretty compelling, right?
- Savage nitpicking. Next you learn to ask methodological questions: why did the authors omit that control, or choose this particular method of statistical analysis? Why do the experiments in this order and not that order? Suddenly every paper looks like complete crap.
- Understanding. You begin to understand what is good and what is bad about a paper, and how it fits into the context of the field. This does require some familiarity with that context, but it's also a matter of general experience.
I think I'm more or less at the Credulity stage. I can figure out what a paper is about, though it takes me a while, and I get super excited about the key results. I've gotten to the point where I can read papers by myself, but I've never seen a paper picked apart, analyzed, and criticized in any detail where I could actually follow the conversation (this is hard in high school). So I think I'll be needing professional help, as it were, to move beyond Credulity.
(I might have gone a little bit overboard with the paper-reading classes for this semester, though. I added it up and I think I'm going to be reading 6-8 papers a week, or about one per day. Wow.)
Monday, February 8, 2010
Courses for this semester
This semester's coursework has two themes: "Let's Read And Discuss Primary Literature", and "Time To Stop Abstracting Away The Physical Nature Of Biology". Both of my required bioengineering core classes are about cells, molecules, and tissues from a mechanical engineer's perspective. All three of my electives are literature-discussion classes on various topics.
I plan to blog a lot more about the papers I read in these classes. It probably won't be a full Journal Club post for every paper because I'll be reading about 6-8 papers a week, but I'll at least try to summarize them. (This is partly for your benefit and partly for mine -- I expect that writing paper summaries for the blog will help me read and understand the papers better.) But I will do full Journal Club posts for the papers that I find most interesting, and definitely for the ones I present. I might also write about interesting things that happen in the other two classes.
- 20.310 Molecular, Cellular, and Tissue Biomechanics - think introductory mechanics, only all the examples are biological things instead of, you know, steel beams or something. Up till now, I've been accustomed to thinking of DNA as a string of digital (AGCT) information, or maybe as a helical molecule; now it's time to think of DNA as a charged elastic rod, or a randomly-walking polymer. How much force can a motor protein exert to pull a vesicle where it's going? What happens when you push and pull on the cytoskeleton? What's the effect of pressure on cells? How do bones reshape themselves in response to forces?
- 20.330 Fields, Forces, and Flows in Biological Systems - fluids and E&M, only again all the examples are biological things. How do things like diffusion and electrophoresis work? How can you model the cell membrane as an RC circuit? What's the best shape / flow pattern for this sample chamber so that the most protein binds to the sensor on one side?
- 20.385 Advanced Topics in Synthetic Biology - this is paired with a freshman design/seminar course, 20.20, which I took two years ago and which rocked my world so hard. It's a great introduction to synthetic biology. The frosh get to do design projects, and because it's surprisingly hard to do this when you've only had introductory biology, the upperclassmen mentor the frosh teams. And when we're not busy mentoring, we have synthetic biology journal club. I'll be presenting a couple of papers and I'm super pumped.
- 7.25 Biological Regulatory Mechanisms - this just sounds fascinating. All the different ways gene expression or protein action can be controlled. Apart from being cool, this is also highly relevant for synthetic biology. Even apart from that, I'm excited about the lectures. This class involves picking apart the experimental logic of papers in a more rigorous way than I've ever had before, which I'm sure will be good for me as well as being fun. We're focusing on demonstrating results and excluding alternative explanations to an extent that seems to be missing in the modern age of "let's generate a zillion data points and then sift through them". Plus, it's a chance to pick the brains of some aged, sage professors. All in all, it really reminds me of ((my interpretation of) what Raffi said about) learning Talmud.
- 7.346 RNAi: A Revolution in Biology and Therapeutics - yet another paper reading class. I know nothing at all about RNA interference, but it's extremely important both theoretically and (potentially) medically. I'm also interested in using it to make synthetic-biological parts that don't crosstalk as much as protein-based parts do; we'll see if that's feasible.
I plan to blog a lot more about the papers I read in these classes. It probably won't be a full Journal Club post for every paper because I'll be reading about 6-8 papers a week, but I'll at least try to summarize them. (This is partly for your benefit and partly for mine -- I expect that writing paper summaries for the blog will help me read and understand the papers better.) But I will do full Journal Club posts for the papers that I find most interesting, and definitely for the ones I present. I might also write about interesting things that happen in the other two classes.
Monday, February 1, 2010
Academic identity crisis
My dad and I happened to be talking about construction failures earlier today -- a raised highway section that collapsed in the Loma Prieta earthquake; the tiles that fell off the Big Dig ceiling. He spoke about them in terms of redundancy and single points of failure, using the engineer lingo that he's picked up from reading books about civil engineering. When I'm being cynical about synthetic biology, I think biologists like to use these terms to make themselves sound sophisticated.
It does bother me, though, that I will graduate from this place with the word "Engineering" on my degree but possibly without the ability to analyze a design, find its flaws, and fix them. I will be able to design an experiment to determine whether Protein X affects Process Y in the cells of Species Z, which covers the word "Biological" on my degree... but will I be a real engineer?
A lot of synthetic biologists come into the field from computer science or electrical or civil engineering, and the whole point of synthetic biology is to turn biology into a Real Engineering Discipline that systematically uses ideas like redundancy in design. If you count up the most prominent synthetic biologists and their origins, it's easy to get the idea that "foreigners" from Real Engineering Disciplines are coming into biology, understanding the science within a week or two, and then dragging it kicking and screaming out of laboratories into design studios and factories. This, too, bothers me.
I do believe that this endeavor will be a truly collaborative one, and will require people with a thorough grounding in biology-the-science as well as people with classical engineering training. I even have a couple of professors' opinions to back this up, although I won't get actual data to confirm or deny this belief until I'm at least in grad school. Certainly biological training helps with designing and conducting experiments; you have to know what a Western blot does in order to know when it's appropriate to do one, and to do it properly. But this makes the biologists sound like the servant underclass in the making of synthetic biology.
Even apart from all this, I worry that synthetic biology, as the newest addition to the family of bioengineering subfields, is not yet ready for its practitioners to be trained in bioengineering instead of in biology or in engineering. I worry that, with an interdisciplinary education, the only thing I'm becoming good at is dabbling, and that I won't develop a true expertise in any particular subfield. -- Then again, isn't undergrad for exploring, and grad school for developing a focus?
It does bother me, though, that I will graduate from this place with the word "Engineering" on my degree but possibly without the ability to analyze a design, find its flaws, and fix them. I will be able to design an experiment to determine whether Protein X affects Process Y in the cells of Species Z, which covers the word "Biological" on my degree... but will I be a real engineer?
A lot of synthetic biologists come into the field from computer science or electrical or civil engineering, and the whole point of synthetic biology is to turn biology into a Real Engineering Discipline that systematically uses ideas like redundancy in design. If you count up the most prominent synthetic biologists and their origins, it's easy to get the idea that "foreigners" from Real Engineering Disciplines are coming into biology, understanding the science within a week or two, and then dragging it kicking and screaming out of laboratories into design studios and factories. This, too, bothers me.
I do believe that this endeavor will be a truly collaborative one, and will require people with a thorough grounding in biology-the-science as well as people with classical engineering training. I even have a couple of professors' opinions to back this up, although I won't get actual data to confirm or deny this belief until I'm at least in grad school. Certainly biological training helps with designing and conducting experiments; you have to know what a Western blot does in order to know when it's appropriate to do one, and to do it properly. But this makes the biologists sound like the servant underclass in the making of synthetic biology.
Even apart from all this, I worry that synthetic biology, as the newest addition to the family of bioengineering subfields, is not yet ready for its practitioners to be trained in bioengineering instead of in biology or in engineering. I worry that, with an interdisciplinary education, the only thing I'm becoming good at is dabbling, and that I won't develop a true expertise in any particular subfield. -- Then again, isn't undergrad for exploring, and grad school for developing a focus?
Saturday, January 23, 2010
You can talk to me! Can I talk to you?
Hi everyone! This is Paul from Colorado. He's a masters student in Course 16 (Aero-Astro), and he's in the Air Force. He's currently taking this really awesome sounding class called System Architecture (ESD.34), and his project is to analyze this really neat-sounding military helicopter. The idea of the class, and of the project, is that you take a sort of half-reversed approach to analyzing complex engineered systems. What is this part for? How does it relate to the other parts around it, and how does it fit into the system as a whole? How did this system evolve? What was the designer thinking at this step? How can I take this big monolithic-looking complicated thing and break it up into meaningful parts? The helicopter he's looking at is an 80s model which was based on an architecture originally built in the 60s for search and rescue, then adapted for some of the surprisingly similar circumstances of special ops (flying at night in horrible weather, for example), but with the greatly expanded capabilities necessary to deal with things like hostile airspace.
Most other people in his class are also analyzing large mechanical devices (one girl is doing the ISS), but one or two are doing more abstract things. Someone is studying air traffic control, and oh there was another neat project topic that I unfortunately forgot.
We thought it would be amusing to analyze the Saferide shuttle system from this perspective. If it hadn't inexplicably held up for ten minutes at the student center, we would never have started talking.
 Figure 1: No, really.
Figure 1: No, really.
Why don't I do this more often? Just strike up a conversation with random people? You do have to exercise care in choosing the right opening question ("What are you doing this glorious IAP?") and refrain from talking too much about yourself... but it really, really isn't hard. There's not even any risk. The worst that happens is that the conversation fizzles and you go back to your book or iPod or whatever. But despite what people tend to think when locked up inside their own heads, everyone else is a thinking human being with interests and passions and opinions and experiences, and these can differ from your own in fascinating ways.
I'm not saying that everyone should be engaged in conversation all the time. I enjoy reading on the bus too, and I certainly enjoy solitary time, being somewhat introverted. But there's no reason to fear or avoid the nontrivial stranger conversation.
Tomorrow I think I'll strike up a conversation with the bus driver.
[The only problem I have with the "Just Shy" shirt is that it automatically puts the onus on the other party to initiate conversation. I think that, in wearing it, one must also resolve to initiate more conversations oneself. If that is done, there can be no harm in advertising that your ears are open for business.]
Most other people in his class are also analyzing large mechanical devices (one girl is doing the ISS), but one or two are doing more abstract things. Someone is studying air traffic control, and oh there was another neat project topic that I unfortunately forgot.
We thought it would be amusing to analyze the Saferide shuttle system from this perspective. If it hadn't inexplicably held up for ten minutes at the student center, we would never have started talking.
Figure 1: No, really.Why don't I do this more often? Just strike up a conversation with random people? You do have to exercise care in choosing the right opening question ("What are you doing this glorious IAP?") and refrain from talking too much about yourself... but it really, really isn't hard. There's not even any risk. The worst that happens is that the conversation fizzles and you go back to your book or iPod or whatever. But despite what people tend to think when locked up inside their own heads, everyone else is a thinking human being with interests and passions and opinions and experiences, and these can differ from your own in fascinating ways.
I'm not saying that everyone should be engaged in conversation all the time. I enjoy reading on the bus too, and I certainly enjoy solitary time, being somewhat introverted. But there's no reason to fear or avoid the nontrivial stranger conversation.
Tomorrow I think I'll strike up a conversation with the bus driver.
[The only problem I have with the "Just Shy" shirt is that it automatically puts the onus on the other party to initiate conversation. I think that, in wearing it, one must also resolve to initiate more conversations oneself. If that is done, there can be no harm in advertising that your ears are open for business.]
Subscribe to:
Posts (Atom)

{kind=link}
{kind=link}