But I'm still not in the habit of putting semicolons after everything. Despite studying Java and Scheme in high school, I never got very good with them or did anything practical. Right now, I'd say my native tongue is Python. Maybe if I get into automation, one of these days I'll
Monday, October 18, 2010
Chasing down particles
I'm up late writing MATLAB code to analyze a whole load of particle tracking data. It's not exactly a fun process, but I can feel myself getting better at MATLAB. It's like the coding equivalent of eating all my asparagus. There are some great tricks you can do to slice and dice arrays with fancy indexing. But, ugh, the syntax for optional arguments makes me wince (and avoid using optional arguments, unfortunately).
But I'm still not in the habit of putting semicolons after everything. Despite studying Java and Scheme in high school, I never got very good with them or did anything practical. Right now, I'd say my native tongue is Python. Maybe if I get into automation, one of these days I'llclamber up on the big-programmer potty learn C/C++.
But I'm still not in the habit of putting semicolons after everything. Despite studying Java and Scheme in high school, I never got very good with them or did anything practical. Right now, I'd say my native tongue is Python. Maybe if I get into automation, one of these days I'll
Monday, October 11, 2010
Storycest Is Best
For my folk song class, I'm writing an analysis of the old Scottish ballad Kemp Owyne (specifically the version called Kempion; here are some related ballads*). The most interesting part, I find, is looking at the relationships between the different versions and between Kemp Owyne and other stories.
Discovery number one: no version of Kemp Owyne is 'complete' in the sense of containing all the plot elements found in any version of Kemp Owyne. Furthermore, the different versions ('subspecies'?) seem to differ not randomly but systematically in the elements they omit. The Kempion version leaves out the vast majority of the beginning, so for example, you never find out why the evil stepmother curses the princess (in fact, you don't even find out it was the evil stepmother's fault until the very end), whereas other versions start out with a thread where the stepmother is angry about not being called "the fairest of them all". It's hard for me to even imagine what it's like to only know one version of the story, since I'm in the privileged position of being able to read them all and cross-reference.
Discovery number two: there's a lot more swapping of story elements than I thought -- a lot more random ligation of characters into new situations. I suppose most folktales are fanfiction of other folktales, for some sense of 'fanfiction'. For example, Kemp Owyne is supposed to be identified with Ywain/Yvain from the Arthurian legends, aka the historical Owain mab Urien (and "Kemp" means 'hero' or 'champion'). Why? No reason. He's just a convenient fictional knightly hero type. But interestingly enough, there's an invocation to St. Mungo at the end of Kempion... and yet, Yvain is also supposedly St. Mungo's father! How did that happen?? Either there is massive storycest going on, or someone chose to invoke a saint who wasn't even born yet. (You have to admit that time-traveling Scots heroes and saints would be pretty awesome.)
It really is like evolution, complete with horizontal gene transfer. But also with revival and retelling and all sorts of processes that don't have obvious biological analogues. Fascinating!
[*] Those texts have some typos in them, but I wasn't about to type all of Kemp Owyne again, and I'm really fond of that whole website because it makes up in quantity what little it lacks in quality. (The typos are few enough that I feel like only scholars really ought to care about them.)
Discovery number one: no version of Kemp Owyne is 'complete' in the sense of containing all the plot elements found in any version of Kemp Owyne. Furthermore, the different versions ('subspecies'?) seem to differ not randomly but systematically in the elements they omit. The Kempion version leaves out the vast majority of the beginning, so for example, you never find out why the evil stepmother curses the princess (in fact, you don't even find out it was the evil stepmother's fault until the very end), whereas other versions start out with a thread where the stepmother is angry about not being called "the fairest of them all". It's hard for me to even imagine what it's like to only know one version of the story, since I'm in the privileged position of being able to read them all and cross-reference.
Discovery number two: there's a lot more swapping of story elements than I thought -- a lot more random ligation of characters into new situations. I suppose most folktales are fanfiction of other folktales, for some sense of 'fanfiction'. For example, Kemp Owyne is supposed to be identified with Ywain/Yvain from the Arthurian legends, aka the historical Owain mab Urien (and "Kemp" means 'hero' or 'champion'). Why? No reason. He's just a convenient fictional knightly hero type. But interestingly enough, there's an invocation to St. Mungo at the end of Kempion... and yet, Yvain is also supposedly St. Mungo's father! How did that happen?? Either there is massive storycest going on, or someone chose to invoke a saint who wasn't even born yet. (You have to admit that time-traveling Scots heroes and saints would be pretty awesome.)
It really is like evolution, complete with horizontal gene transfer. But also with revival and retelling and all sorts of processes that don't have obvious biological analogues. Fascinating!
[*] Those texts have some typos in them, but I wasn't about to type all of Kemp Owyne again, and I'm really fond of that whole website because it makes up in quantity what little it lacks in quality. (The typos are few enough that I feel like only scholars really ought to care about them.)
Sunday, September 26, 2010
Science poetry: Heredity, Thomas Hardy
I am the family face;
Flesh perishes, I live on,
Projecting trait and trace
Through time to times anon,
And leaping from place to place
Over oblivion.
The years-heired feature that can
In curve and voice and eye
Despise the human span
Of durance -- that is I;
The eternal thing in man,
That heeds no call to die
--Thomas Hardy [source]
I like the alliterations and internal-half-rhymes in the first stanza. They remind me of what little I know about Anglo-Saxon verse. (I read Beowulf once...)
Monday, September 13, 2010

Sunday, September 5, 2010
$18 can buy a lot of fun
(My living group is doing Rush right now, which means my life is basically eaten for the next two weeks, such that I feel no shame about writing a punt-tastic post like this.)
I will just note that $18 can buy more dry ice than twenty MIT students can play around with for three hours. Try it out some time. Make sure to bring soapy water, interestingly-shaped glass vessels, small white and colored lights/LEDs, coins, spoons, and hot water.
Try putting a little water in a spoon and then resting the spoon on top of a chunk of dry ice. You can see the water freeze right before your eyes. It's pretty cool.
I will just note that $18 can buy more dry ice than twenty MIT students can play around with for three hours. Try it out some time. Make sure to bring soapy water, interestingly-shaped glass vessels, small white and colored lights/LEDs, coins, spoons, and hot water.
Try putting a little water in a spoon and then resting the spoon on top of a chunk of dry ice. You can see the water freeze right before your eyes. It's pretty cool.
Sunday, August 29, 2010
What's wrong with the cartoon eukaryotic cell? [Unusual Cells pt. 1]
I'm starting a series of posts based on a class I taught about "Unusual Cells" for Splash. Eventually, each post will include links to all the others.
The point of this series is to understand the wild and crazy unusual cells that populate the world (and that populate us!). But we should first understand the usual cell... or, perhaps, we will find that there is no such thing as a usual cell.
Consider the cartoon eukaryotic cell that we all know and, presumably, love:
 Figure 1: Typical eukaryotic cell. I'll refrain from listing the organelles, in order to prevent yawning; if you're curious, check Wikipedia, where this image is from.
Figure 1: Typical eukaryotic cell. I'll refrain from listing the organelles, in order to prevent yawning; if you're curious, check Wikipedia, where this image is from.
So... what's wrong with this cell? Here's a sampling of some of the answers my Splash students have given me:
The answer I'm really looking for is this:
(To be fair, Splash students pretty much always get this one too, and it's a general case of the answers I gave above except for #3.)
There's nothing in this cell! Yes, the major organelles are there, but where is the cytoskeleton? It's just that tiny little fiber (#7), which you can barely even see. Real cells are just packed with stuff. Structural proteins, membrane proteins, highways of vesicles wandering to and fro...
Now, of course, this is kind of a necessary evil. We can't include all that stuff in cartoons of the cell that are supposed to show the major organelles, because it would just be a distraction. Visual noise. For example, check out this slice of a more realistic watercolor:
 Figure 2: You can see a bit of the Golgi (yellow stacks) in this picture. The geodesic-dome-looking thing is a protein framework that's making a vesicle bud out from the Golgi. [Source]
Figure 2: You can see a bit of the Golgi (yellow stacks) in this picture. The geodesic-dome-looking thing is a protein framework that's making a vesicle bud out from the Golgi. [Source]
Seriously, check out the whole thing. It's beautiful. If I could get a quality print of this I would hang it above my bed. But even this is far from showing everything. The empty space between all those blobs is filled with crazy amounts of ions, small molecules, and of course water.
For a different perspective, check out these photos of cells in which the cytoskeleton has been labeled with green fluorescence. Yep, that's just the cytoskeleton... it reaches everywhere, helping the cell maintain its shape and move around (just like the human skeleton), and giving direction to packages of important chemicals as they motor their way hither and thither (something like the human circulatory system).
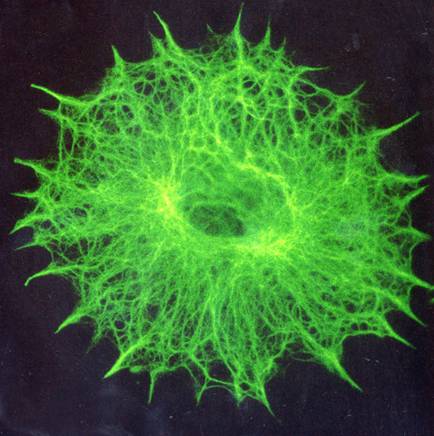

To do: find out what percentage of the membrane surface area is proteins. I know this is in one of my textbooks somewhere.
The point of this series is to understand the wild and crazy unusual cells that populate the world (and that populate us!). But we should first understand the usual cell... or, perhaps, we will find that there is no such thing as a usual cell.
Consider the cartoon eukaryotic cell that we all know and, presumably, love:
Figure 1: Typical eukaryotic cell. I'll refrain from listing the organelles, in order to prevent yawning; if you're curious, check Wikipedia, where this image is from.So... what's wrong with this cell? Here's a sampling of some of the answers my Splash students have given me:
- You can't see the DNA
- You can't see the proteins
- It's cut in half (yes, some of them are smartasses)
- There are no membrane proteins
The answer I'm really looking for is this:
It's empty!
(To be fair, Splash students pretty much always get this one too, and it's a general case of the answers I gave above except for #3.)
There's nothing in this cell! Yes, the major organelles are there, but where is the cytoskeleton? It's just that tiny little fiber (#7), which you can barely even see. Real cells are just packed with stuff. Structural proteins, membrane proteins, highways of vesicles wandering to and fro...
Now, of course, this is kind of a necessary evil. We can't include all that stuff in cartoons of the cell that are supposed to show the major organelles, because it would just be a distraction. Visual noise. For example, check out this slice of a more realistic watercolor:
Figure 2: You can see a bit of the Golgi (yellow stacks) in this picture. The geodesic-dome-looking thing is a protein framework that's making a vesicle bud out from the Golgi. [Source]Seriously, check out the whole thing. It's beautiful. If I could get a quality print of this I would hang it above my bed. But even this is far from showing everything. The empty space between all those blobs is filled with crazy amounts of ions, small molecules, and of course water.
For a different perspective, check out these photos of cells in which the cytoskeleton has been labeled with green fluorescence. Yep, that's just the cytoskeleton... it reaches everywhere, helping the cell maintain its shape and move around (just like the human skeleton), and giving direction to packages of important chemicals as they motor their way hither and thither (something like the human circulatory system).
To do: find out what percentage of the membrane surface area is proteins. I know this is in one of my textbooks somewhere.
Monday, August 23, 2010
Perhaps I should flip a coin?
I'm working out my class schedule for the upcoming semester, and I've run into a bit of a dilemma. Two classes are at the exact same time. Both clamor for my attention. Both are only offered in the fall, and this is my last year. They are as mutually exclusive as it is possible for two classes to be.
THE CANDIDATES
THE ARGUMENT
I want to study systems biology in grad school. Therefore, I should get started. Taking this class may help me with my continuing work in the Silver lab, and might even help me get into a good grad program.
THE COUNTERARGUMENT
I will have plenty of time to study systems biology in grad school. (And if I really get an itch, I can always pick up Uri Alon's book.) I should take this chance to explore a humanities topic that I'm really interested in, while I'm still an undergrad, because time is short.
------------------------------------------
So, what do I do? Both of these arguments are fairly convincing to me. Which one wins? Or, are there other arguments I've missed?
THE CANDIDATES
- 7.32 Systems Biology: This field is a sister to synthetic biology. I'm interested in to the point of wanting to pursue it in grad school. The Silver lab, where I'm working, is a systems biology lab in more than just name (although it's certainly not typical, being focused on engineering). Networks! Switches! Stochastic behavior! Dynamics! Oscillators! Pattern formation!
- 21A.212 Myth, Religion, and Symbolism: This class looks like it's going to hit one of my biggest avocational buttons. Despite being atheist/agnostic/nonreligious (damn labels), I've always had a fascination with the power of ritual and storytelling -- the roles they play in our lives and how they adapt to non-religious contexts. How did I manage to not notice this class existed before?
THE ARGUMENT
I want to study systems biology in grad school. Therefore, I should get started. Taking this class may help me with my continuing work in the Silver lab, and might even help me get into a good grad program.
THE COUNTERARGUMENT
I will have plenty of time to study systems biology in grad school. (And if I really get an itch, I can always pick up Uri Alon's book.) I should take this chance to explore a humanities topic that I'm really interested in, while I'm still an undergrad, because time is short.
------------------------------------------
So, what do I do? Both of these arguments are fairly convincing to me. Which one wins? Or, are there other arguments I've missed?
Subscribe to:
Posts (Atom)
