Typical. I don't get an idea for an Ada Lovelace Day post until well after the day itself has passed. "But I just don't know any women scientists other than the obvious ones!" Not an excuse. How many women scientists was I taught by in elementary and middle school, completely oblivious to their prior lives? Of course, I was taught by men scientists too, and they have equally interesting stories. I'm just going to focus on one particular woman for the moment.
Molly was the science specialist at my elementary school. She gave the impression of a powerful but friendly science warlock, introducing us grade-school apprentices to a world of wonderful things. Yes, it sounds cheesy, but there you are. I don't have any data, but I think I must credit Molly with giving me that initial (strong) inclination toward science that tipped and boiled over when I ran into a copy of The Cartoon Guide to Genetics (Gonick & Wheelis). It's been more than a decade since then, and I've never looked back.
I can only remember a few fragments of specific things that Molly taught us, but I distinctly remember some of the things that we did. My school had a lot of forested property around the buildings, some of which had been invaded by the vinca that someone decided to plant around the parking lots. We spent many an hour tramping around the woods, of course, and not only identifying trees and noticing erosion, but waging war on the vinca. We marked out test plots and poured mulch over some, staked black plastic over others, and wielded picks, shovels, and good ol' pulling on the rest. I lost track of the project once I was in middle school, so I don't know how successful the ongoing efforts were, but it was pretty awesome at the time.
We also -- this is kind of surprising, in retrospect -- had a strong focus on communication. At some level, every third grader is going to remember "communicating" (writing the occasional couple of paragraphs). But we wrote full-length letters to lawmakers about environmental issues (not skimping on either the science or the rhetoric), and drew a cartoon illustrating a forest fire. The final exhibition of everyone's cartoons was a sight to see.
The other day, I happened to idly wonder what Molly was up to now, and so I googled her. I found out little about what she's up to since her teaching career, but there was quite a bit about what she had been up to before taking up the overhead projector. She published several papers in the sixties about Blepharisma and the recovery of cells from X-ray irradiation. She was at Stanford! Doing real science! Back in the day, before sequencing and so on were invented! Why was I not informed?
She also had an active life outside school, running marathons and working with the San Francisco Sidewalk Astronomers to build DIY Dobsonian telescopes. Man, how cool is that?
Molly Lusignan, scientist, scholar, teacher, and all-around awesome woman, I salute you. The moral of the story for the rest of us is to find out what our teachers (and our children's teachers) do outside of the classroom, about their history, their hobbies, and their dreams. These days, it's of course made easy to stalk people via Google (and Google Scholar! <3), but there's a lot more to be said for connecting in person. I wish I could have found her current contact information...
Monday, March 29, 2010
Sunday, March 21, 2010
Journal Club Followup: The amber-suppressing AND gate
We went over the AND gate paper in class, and generated a lot of constructive criticism that I hadn't thought of by just reading it on my own. I guess this is what class is for, huh.
In particular, we addressed the question of whether the AND gate is truly modular. As I discussed in the previous post, in principle this AND gate is modular in the sense that it can be plugged into different inputs and outputs. However, this isn't the whole story. "Plugging it in" to a different input is not so simple as just putting a new promoter in front of Input Gene 1. The whole promoter/RBS/coding-region assembly has to be tuned to have an appropriate strength. If you swap in a stronger promoter, you might have to weaken the RBS (ribosome binding site) in order to get just the right amount of protein expression to feed in to the rest of the AND gate.
In fact, that's exactly what the authors had to do when they first put their AND gate together, which I kind of glossed over. Recall that the system works by expressing an mRNA with amber stop codons in, and the amber-suppressor tRNA that can read those stop codons; the mRNA codes for a protein that transcribes the output promoter. Now, in principle this should Just Work. But in practice, if there's enough mRNA around, you can get spontaneous read-through even if the amber-suppressor tRNA is supposedly turned "off", for two reasons. One, even if the tRNA is "off", it might still be produced at a low basal level. Two, the amber stop codon is relatively "weak", and sometimes just gets read through anyway. (That is, it's not very good at recruiting the translation-stopping machinery, which is supposed to disassemble the ribosome and cut loose the newly translated protein.) So the authors had to adjust the RBS that governed translation of the mRNA, so there wouldn't be too much of it floating around and you wouldn't get this spurious effect.
Apparently this sort of adjustment is called "impedance matching", for those of you who are more familiar with electronics than cells. I don't know enough about electronics to explain exactly what impedance matching is or why it's a suitable analogy here, but it boils down to "make sure all the wires are carrying appropriate amounts of current, and if you connect something new to a wire you might have to add a resistor or something to fix the current back to how it used to be."
So, no, it's not plug-and-play quite yet. More like plug, mutagenize the RBS, and play... but maybe we'll get there eventually.
The other main criticism of this AND gate, from a modularity point of view, is that you can't have two copies of the gate in the same cell and expect them to operate independently. mRNAs and tRNAs float around, and if one AND gate is expressing mRNA and the other is expressing tRNA, then both of them will output ON, even though both of them ought to be OFF. This is a more serious problem, because you can't just tune an RBS and expect it to go away -- this design for an AND gate is in principle not modular with respect to other AND gates placed in the same cell. (Possible workarounds include hiding different AND gates in different cells and mixing several populations together, but then you have to work with cell-to-cell cooperation, which is a whole different ballgame.)
In particular, we addressed the question of whether the AND gate is truly modular. As I discussed in the previous post, in principle this AND gate is modular in the sense that it can be plugged into different inputs and outputs. However, this isn't the whole story. "Plugging it in" to a different input is not so simple as just putting a new promoter in front of Input Gene 1. The whole promoter/RBS/coding-region assembly has to be tuned to have an appropriate strength. If you swap in a stronger promoter, you might have to weaken the RBS (ribosome binding site) in order to get just the right amount of protein expression to feed in to the rest of the AND gate.
In fact, that's exactly what the authors had to do when they first put their AND gate together, which I kind of glossed over. Recall that the system works by expressing an mRNA with amber stop codons in, and the amber-suppressor tRNA that can read those stop codons; the mRNA codes for a protein that transcribes the output promoter. Now, in principle this should Just Work. But in practice, if there's enough mRNA around, you can get spontaneous read-through even if the amber-suppressor tRNA is supposedly turned "off", for two reasons. One, even if the tRNA is "off", it might still be produced at a low basal level. Two, the amber stop codon is relatively "weak", and sometimes just gets read through anyway. (That is, it's not very good at recruiting the translation-stopping machinery, which is supposed to disassemble the ribosome and cut loose the newly translated protein.) So the authors had to adjust the RBS that governed translation of the mRNA, so there wouldn't be too much of it floating around and you wouldn't get this spurious effect.
Apparently this sort of adjustment is called "impedance matching", for those of you who are more familiar with electronics than cells. I don't know enough about electronics to explain exactly what impedance matching is or why it's a suitable analogy here, but it boils down to "make sure all the wires are carrying appropriate amounts of current, and if you connect something new to a wire you might have to add a resistor or something to fix the current back to how it used to be."
So, no, it's not plug-and-play quite yet. More like plug, mutagenize the RBS, and play... but maybe we'll get there eventually.
The other main criticism of this AND gate, from a modularity point of view, is that you can't have two copies of the gate in the same cell and expect them to operate independently. mRNAs and tRNAs float around, and if one AND gate is expressing mRNA and the other is expressing tRNA, then both of them will output ON, even though both of them ought to be OFF. This is a more serious problem, because you can't just tune an RBS and expect it to go away -- this design for an AND gate is in principle not modular with respect to other AND gates placed in the same cell. (Possible workarounds include hiding different AND gates in different cells and mixing several populations together, but then you have to work with cell-to-cell cooperation, which is a whole different ballgame.)
Tuesday, March 9, 2010
Where I'm going to be this summer
I hope you can forgive me for prematurely tooting my own horn, as there are many logistical details still to be worked out, but... I'm going to be working in Pam Silver's lab this summer! Squee!
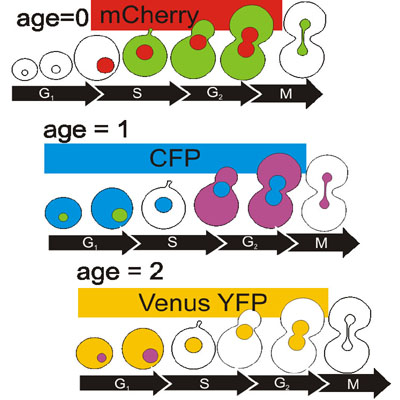 Figure 1: Yeast cell cycle counters! Hmm, those sound awfully familiar....
Figure 1: Yeast cell cycle counters! Hmm, those sound awfully familiar....
Figure 1: Yeast cell cycle counters! Hmm, those sound awfully familiar....Saturday, March 6, 2010
The answer to my question about scaffolds
I passed my scaffold question on to my 20.20 professors, and got a very thoughtful response. (To their credit, it was also a very timely response; I've just been slow in posting it.)
Recall my question: there seemed to be a conflict between the role of kinase cascades as signal amplifiers, and the role of scaffolds as giving specificity. Turns out, naturally, that there isn't really a conflict at all. Rather, I didn't fully understand what scaffolds do. The crux of what I wasn't understanding is that proteins go on and off their scaffold all the time -- they don't stay bound (semi)permanently.
One of the main benefits of scaffolds is to make signaling pathways more efficient: they help bring proteins close together so they can interact. If you have only a few molecules of Protein A and Protein B, then they may not encounter each other very often if they're just freely diffusing around the cell. A scaffold can grab one Protein A and one Protein B, hold them close together long enough to interact, and the release them and go look for two new proteins. Or if A is a kinase for B, then A can stay on the scaffold and multiple copies of B can take turns.
Scaffolds also act like little switchboards, dictating where signaling pathways are allowed to cross and merge or where they must stay separate. Suppose a scaffold has one binding site for the kinase A, and a second binding site that can fit either Protein B or C, but not D. In this case you get activation of both B and C, and the signal from A propagates in two different directions, provoking two separate responses (but not provoking a third response). This is really helpful because a lot of kinases are promiscuous -- they'll phosphorylate anything they can get their hands on -- so if they're confined to scaffolds, then they'll do whatever the scaffold says they can do, and no more.
Recall my question: there seemed to be a conflict between the role of kinase cascades as signal amplifiers, and the role of scaffolds as giving specificity. Turns out, naturally, that there isn't really a conflict at all. Rather, I didn't fully understand what scaffolds do. The crux of what I wasn't understanding is that proteins go on and off their scaffold all the time -- they don't stay bound (semi)permanently.
One of the main benefits of scaffolds is to make signaling pathways more efficient: they help bring proteins close together so they can interact. If you have only a few molecules of Protein A and Protein B, then they may not encounter each other very often if they're just freely diffusing around the cell. A scaffold can grab one Protein A and one Protein B, hold them close together long enough to interact, and the release them and go look for two new proteins. Or if A is a kinase for B, then A can stay on the scaffold and multiple copies of B can take turns.
Scaffolds also act like little switchboards, dictating where signaling pathways are allowed to cross and merge or where they must stay separate. Suppose a scaffold has one binding site for the kinase A, and a second binding site that can fit either Protein B or C, but not D. In this case you get activation of both B and C, and the signal from A propagates in two different directions, provoking two separate responses (but not provoking a third response). This is really helpful because a lot of kinases are promiscuous -- they'll phosphorylate anything they can get their hands on -- so if they're confined to scaffolds, then they'll do whatever the scaffold says they can do, and no more.
To see the beauty of a paper towel, and the world in an empty dewar
If you ever work in a bio lab, you'll find that often the closest piece of paper to hand is a paper towel, and the closest pen to hand is a sharpie or lab marker. Predictably, a lot of scientific note-taking happens on paper towels. I know I've done my share of calculations on paper towels, and I've heard of some students keeping their notebook as a stack of scribbled-on paper towels. (Shame!)
And then I walk down the infinite corridor and see something like this:
 Figure 1: Yes, Anonymous Lab Worker, someone appreciates your calligraphy. I love it when people do a little to make the world a weirder and more beautiful place. (Click to embiggen; the handwriting actually looks a lot better in high resolution.)
Figure 1: Yes, Anonymous Lab Worker, someone appreciates your calligraphy. I love it when people do a little to make the world a weirder and more beautiful place. (Click to embiggen; the handwriting actually looks a lot better in high resolution.)
And then I walk down the infinite corridor and see something like this:
Monday, March 1, 2010
Ideas dump
I'm looking over some old brainstorms, because of course I'm not busy at all, no sir, ahahaha... Anyway. I keep getting neat ideas, and I'm going to put some of them here so that (1) I don't forget them before I find time to investigate and (2) maybe someone will actually look at them and think about them. Also, I apologize for not explaining these ideas for the benefit of biology neophytes.
- Enzymes that bind DNA and perform actions including cutting/pasting DNA or recruiting other enzymes. Can you inhibit their action by adding short pieces of RNA (or similar) with the same sequence as their binding site?
- RNAi seems to exist in bacteria, kind of (not as much as in C.elegans). Is it useful for creating synthbiological devices? I don't recall seeing any; what's the roadblock?
- In particular, RNAi could be a neat way of getting around the crosstalk/specificity problem, since it's very sequence-specific and designing RNA sequences is easier than designing proteins.
- What would it take to drive localization to a synthetic organelle?
- Need to learn more about riboswitches. All the riboswitches I've seen so far are the kind that respond to a small molecule. Hmm: can you make a riboswitch that responds to a short ssRNA??
- What are the primitives of biological circuits? In a regime where it's easier to build monolithic black boxes than to reuse parts, how do things like two-component signaling evolve, that almost look intelligently designed with modularity etc?
- I think I'm on an RNA kick. Is this justified?
- Whoa, DNA scaffolds? Clever! read this at some point
Which tissues are anisotropic?
Today in my biomechanics class we moved out of molecular mechanics and into tissue mechanics. This will involve a lot of stereotypical mech-E stuff like stress and strain and elasticity that all us biologists have never heard of before -- deterministic, bulk properties of continuous solids (or gooey things), rather than stochastic models of single molecules or long chains of molecules being buffeted around by thermal motion.
In this field, as in every other, you have to make simplifying assumptions. Two of the key assumptions we often make about bulk materials are that they are homogeneous and isotropic. That is, they're uniform throughout, and they're the same in every direction. Tissue like muscle (with fibers) or substances like string cheese (also with fibers) are anisotropic (there's your five dollar word for the day). If you're stretching them and you want to find out how they deform, it matters which direction you're pulling. After we learned what all these fancy words meant, we categorized a few tissues:
Isotropic:
Anisotropic:
As I was writing these down, I noticed a pattern. All the isotropic tissues are what I'd call "biochemical tissues". Their main role is to store chemicals or make reactions happen. In contrast, all the isotropic tissues are "mechanical tissues", that make stuff move or stick together. Huh! Insight! But then it occurred to me that this makes total sense. After all, if you're doing a chemical reaction in a test tube, and you pour it into a bowl, the reaction will still happen. Whereas if you suddenly make all the fibers in a muscle run in a different direction, the muscle will do something completely different. If a tissue's job is to apply a force, it needs to apply that force in the right direction. Biochemical tissues are OK being isotropic, because their job is isotropic, and it seems like it ought to be harder for a developing organism to grow an anisotropic tissue, so why bother.
(Cartilage goes under "isotropic tissues", not because it does much in the way of chemical reactions, but because it's basically just a cushiony substance.)
Neat, huh?
Addendum: what about neurons and nerve tissue?
Well, we can throw out "neurons" straight out, because neurons are single cells, and tissues can only be treated as bulk materials if you've got a lot more than single cells. Trying to treat tissues on the tens-of-micrometers scale as bulk materials is like trying to calculate the viscosity of minestrone. It just doesn't apply. The viscosity of chicken broth is way different from the viscosity of beans. It only works if you zoom way, way out so that the effects of all the little bits and bobs become uniform over the entire blob of whatever you're looking at.
But we can totally look at nerve tissue this way. Your spinal cord, I would imagine, is a little bit like string cheese or rope. It's a whole bunch of long cellular fibers in parallel. If you pulled on it lengthwise, I would expect it to stretch; if you pulled on it widthwise, I would expect it to fray apart. It's anisotropic.
Gray matter in your cortex, on the other hand, I would call isotropic, or at least closer to isotropic (the cortex does have layers after all). Gray matter is mostly made up of cell bodies arranged more or less randomly, not a bunch of fibers all aligned with each other. Having dissected the odd brain or two, I think the best comparison for gray matter might be a firm jelly. (Exactly how firm it is depends on whether it's been preserved and how.)
So, does nerve tissue fit into the "biochemical" vs. "mechanical" tissue dichotomy? Not very cleanly. Then again, it's an atypical tissue. Its job is to send signals from point A to point B. For spinal cords and nerve bundles, point A and point B are far enough apart that it becomes important, on a macro scale, for the cells to go in the right direction. Gray matter contains mostly cell bodies rather than axons, so it's like a pile of Point As (or Points A, for the Captains Pedantic), whose job is to turn chemical signals into electrical activity.
In this field, as in every other, you have to make simplifying assumptions. Two of the key assumptions we often make about bulk materials are that they are homogeneous and isotropic. That is, they're uniform throughout, and they're the same in every direction. Tissue like muscle (with fibers) or substances like string cheese (also with fibers) are anisotropic (there's your five dollar word for the day). If you're stretching them and you want to find out how they deform, it matters which direction you're pulling. After we learned what all these fancy words meant, we categorized a few tissues:
Isotropic:
- Liver
- Fat
- Blood
- Cartilage
Anisotropic:
- Muscle
- Bone
- Skin
- Ligaments
As I was writing these down, I noticed a pattern. All the isotropic tissues are what I'd call "biochemical tissues". Their main role is to store chemicals or make reactions happen. In contrast, all the isotropic tissues are "mechanical tissues", that make stuff move or stick together. Huh! Insight! But then it occurred to me that this makes total sense. After all, if you're doing a chemical reaction in a test tube, and you pour it into a bowl, the reaction will still happen. Whereas if you suddenly make all the fibers in a muscle run in a different direction, the muscle will do something completely different. If a tissue's job is to apply a force, it needs to apply that force in the right direction. Biochemical tissues are OK being isotropic, because their job is isotropic, and it seems like it ought to be harder for a developing organism to grow an anisotropic tissue, so why bother.
(Cartilage goes under "isotropic tissues", not because it does much in the way of chemical reactions, but because it's basically just a cushiony substance.)
Neat, huh?
Addendum: what about neurons and nerve tissue?
Well, we can throw out "neurons" straight out, because neurons are single cells, and tissues can only be treated as bulk materials if you've got a lot more than single cells. Trying to treat tissues on the tens-of-micrometers scale as bulk materials is like trying to calculate the viscosity of minestrone. It just doesn't apply. The viscosity of chicken broth is way different from the viscosity of beans. It only works if you zoom way, way out so that the effects of all the little bits and bobs become uniform over the entire blob of whatever you're looking at.
But we can totally look at nerve tissue this way. Your spinal cord, I would imagine, is a little bit like string cheese or rope. It's a whole bunch of long cellular fibers in parallel. If you pulled on it lengthwise, I would expect it to stretch; if you pulled on it widthwise, I would expect it to fray apart. It's anisotropic.
Gray matter in your cortex, on the other hand, I would call isotropic, or at least closer to isotropic (the cortex does have layers after all). Gray matter is mostly made up of cell bodies arranged more or less randomly, not a bunch of fibers all aligned with each other. Having dissected the odd brain or two, I think the best comparison for gray matter might be a firm jelly. (Exactly how firm it is depends on whether it's been preserved and how.)
So, does nerve tissue fit into the "biochemical" vs. "mechanical" tissue dichotomy? Not very cleanly. Then again, it's an atypical tissue. Its job is to send signals from point A to point B. For spinal cords and nerve bundles, point A and point B are far enough apart that it becomes important, on a macro scale, for the cells to go in the right direction. Gray matter contains mostly cell bodies rather than axons, so it's like a pile of Point As (or Points A, for the Captains Pedantic), whose job is to turn chemical signals into electrical activity.
Subscribe to:
Posts (Atom)

{kind=link}